Категории
Категории Корзина
Корзина Информация
Информация Хиты продаж
Хиты продаж






Особенности строения зеленых водорослей
особенности строения, представители вида и их количество, основное предназначение
Зеленые водоросли представляют собой обширнейшую группу живых микроорганизмов, ткани которых практически не подлежат какой-либо дифференциации, поэтому их по-прежнему относят к группе низших растений, включающих в себя, помимо обозначенного вида, бактерии, лишайники и грибы.
 Зеленые водоросли считаются низшими организмами на ряду с бактериями и грибами
Зеленые водоросли считаются низшими организмами на ряду с бактериями и грибамиБиологический вид Chlorophyta
Отдел Chlorophyta включает в себя огромный список зеленых водорослей, который по предварительным оценкам ученых-исследователей исчисляется тысячами видов растений (приблизительное их число варьируется в крайне широком диапазоне от 13 до 20 тысяч).
Именно поэтому определить точное количество их видов на сегодняшний день невозможно и главная причина такого положения заключается в сложности дифференциации этих растительных организмов.
Представители отдела
Практически все виды таллома можно встретить среди зеленых водорослей. Представителями этого отдела являются не только простейшие одноклеточные и многоклеточные колониальные водоросли планктонного типа, неспособные сопротивляться течению водных масс, но и одно- и многоклеточный фитобентос, отличительная черта которого заключается в его среде обитания — океанском, морском или речном грунте.
 Не все представители класса зеленых водорослей имеют зеленый цвет
Не все представители класса зеленых водорослей имеют зеленый цветЕдинственным исключением являются одно- и многоклеточные растения ризоподиального типа с их сложной структурой строения и отличным талломом.
Тем не менее именно рассматриваемый отдел официально признан самым обширным во всем растительном мире водоемов. Несмотря на название, далеко не все представители обладают слоевищем (телом) зеленого оттенка. К примеру, такой представитель вида, как трентеполия, имеет клетки оранжево-красного пигмента, обусловленные накоплением каротина и всевозможных производных этого полезного вещества.
Встречаются среди этих водорослей и растения нитчатого типа, главное отличие которых сводится не к преобладающему пигменту другого цвета, а к среде обитания. Если в начале своего жизненного пути их можно отнести к бентосам, так как нитчатые водоросли имеют свойство крепиться к водоемному грунту, то со временем они пускаются в свободное плавание, фактически становясь планктонными водорослями, формирующими многочисленные шары и маты из своих колоний.
Классы зеленых водорослей
Из-за сложности идентификации талломов растительных организмов, входящих в отдел водорослей зеленого цвета, а также их более чем внушительного количества, ученым до сих пор так и не удалось создать какой-то единый их реестр или подробный классификатор. Тем не менее в настоящее время выделяют пять основных классов зеленых водорослей, к которым относят:
- ульвофициевые;
- брипсодовые;
- хлорофициевые;
- требуксиевые;
- празиновые.
 Представители класса Ulvophyceae обитают преимущественно в морской воде
Представители класса Ulvophyceae обитают преимущественно в морской водеВ класс Ulvophyceae входит не менее тысячи видов растений, преимущественно обитающих в морской среде. Примечательно, что к этому классу относят не только нитчатые и пластинчатые водоросли, но и некоторые виды лишайников. Что же касается морских его представителей, которых абсолютное большинство, то характерной их особенностью является наличие известковых микрочастиц. В свою очередь, этот класс делится на два порядка:
- Улотриксовые, чаще всего обитающие в водоемах с пресной водой, прикрепляясь к каким-либо предметам и образуя многочисленные колонии в виде низкорослых кустиков. Бесполые растения этого порядка имеют свойство размножаться двумя способами, первый из которых подразумевает деление одной нити на несколько, в то время как для второго свойственно образование зооспор в материнских клетках с дальнейшим их выбросом и распространением.
- Ульвовые, которые встречаются во всех морях мира, предпочитая в первую очередь соленые водоемы с теплой водой. Отличительной характеристикой этих водорослей с талломом ярко-зеленого цвета является наличие ножки для крепления к морскому грунту, а также разнообразие строения (слоевище растений может иметь как трубчатую, так и мешковидную или пластинчатую форму).
Класс Bryopsidophyceae насчитывает около пятисот видов растительных организмов, обладающих талломом неклеточного типа. Все эти водоросли относятся к единому порядку с одноименным названием Бриопсидовые, преобладающему в пресных водах. В свою очередь, этот порядок делится на три основных рода: бриопсис, кодиум и каулерпа, отличающихся друг от друга не только строением, но и средой обитания.
Класс одноклеточных водорослей Chlorophyceae насчитывает около 2500 видов, которые относятся к единому порядку вольвоксовых. При этом он делится на два рода: хламидомонада (не менее 500 видов) и вольвокс (около 2 тысяч). Первый род, который также является бесполым, имеет свойство размножаться посредством деления клетки. В то же время вольвокс, который по праву считается наиболее высокоорганизованным представителем этой группы, способен организовывать огромные колонии, насчитывающие тысячи живых организмов.
 Хламидомонада относится к классу зеленых водорослей
Хламидомонада относится к классу зеленых водорослейЕще один класс Trebouxiophyceae состоит из 170 видов одноклеточных организмов, имеющих преимущественно коккоидную форму, которые могут обитать как на земляном грунте, так и на дне пресных озер и рек.
Этот класс классифицируется по двум порядкам на хлорелловые и требуксиевые водоросли, и если первый тип представляет собой неподвижные шарики, для которых естественной средой обитания являются водоемы, то второй входит в состав лишайников, встречающихся на древесной коре либо ведущих свободный образ жизни.
Название последнего класса Prasinophyceae переводится с древнегреческого языка как «зеленый» и на этот раз речь также идет об одноклеточных организмах с коккоидным, жгутиковым или пальмеллоидным типом строения. Делится этот класс на два порядка: пирамимонадовые, которые нередко провоцируют цветение пресных и соленых вод, и хлородендровые подвижного и неподвижного типа со слизкими ножками для крепления к твердым поверхностям. Водоросли последнего порядка часто имеют тот самый красный пигмент из-за наличия каротина. Кроме того, они могут паразитировать, всю жизнь обитая в теле морских червей.
Структура, циклы и предназначение
Несмотря на огромное количество видов зеленых водорослей, которые поражают своим разнообразием, ученые смогли выделить несколько общих характеристик этих живых растительных организмов.
Особенности строения и жизненный цикл
Клетки пластид, которые входят в состав водорослей, чаще всего имеют чашевидную форму, и, помимо классического зеленого, они включают в свой состав желтый (лютеин) и красно-оранжевые пигменты каротиновой группы (зеаксантин, виолаксантин, неоксантин и пр.). Сам хлоропласт имеет особое внутреннее включение — пиреноид, который пронизан 2−6 ограниченными мембранами компартментами, именуемые тилакоидами.
При этом зеленые водорослевые пластиды имеют сдвоенные мембраны, что свойственно красным водорослям и другим более развитым растительным организмам.
 Жизненный цыкл зеленых водорослей зависит от среды их обитания
Жизненный цыкл зеленых водорослей зависит от среды их обитанияКлетки большинства видов рассматриваемой группы имеют сократительные вакуоли, под которыми подразумеваются одномембранные органоиды с клеточным соком в средней части. Не исключено и наличие стигмы — внутриклеточного органоида с ярко-красными пятнами, благодаря которому организм способен двигаться по направлению к или от светового источника.
Практически идентичные по своей структуре жгутики нередко имеют разную длину, при том что их может быть два и более. Эти изоконты не имеют боковых волосков (мастигонем), однако, это вовсе не означает, что у водорослевых жгутиков полностью отсутствуют любые волоски или чешуйки.
 Фактором усиленного роста водорослей является повышение температуры среды их обитания
Фактором усиленного роста водорослей является повышение температуры среды их обитанияЖизненный цикл этих растительных организмов напрямую зависит от их вида и среды обитания. Всего же ученые выделяют три основных цикла их развития:
- Гаплобионтный (зиготическая редукция), для которого свойственно высвобождение гаметы через пору материнской клетки и ее дальнейшее слияние с другой гаметой через трубку. Далее происходит преобразование в зигоспору, в которой после периода покоя образуется 4 зооспоры с последующим формированием полиэдра в каждой из них.
- Гапло-диплобионтный (спорическая редукция), при котором вышедшие из материнской клетки гаметы сливаются в воде, образуя не двух, как в гаплобиотном цикле, а четырехжгутиковые зиготы, свободно парящие в водных массах. Следующий этап предполагает прикрепление зиготы к любой твердой поверхности и ее покрытие плотным слоем, после чего происходит формирование кодиолума, уходящего на длительный покой. Повторная активация этих дубинкообразных клеток, как правило, становится возможной при наступлении благоприятных условий. В итоге из кодиолума выделяется от 4 до 16 спор, формирующих новые нити и все повторяется вновь.
- Диплобионтный (гаметическая редукция), характеризующийся делением крупного ядра, которое образуется при оседании планозиготы и формировании отдельного нитчатого таллома.
Наиболее благоприятным фактором для развития водорослей является повышение температуры, поэтому совсем неудивительно, что их бурный рост можно наблюдать в теплых водах и при наступлении весеннего и летнего сезонов.
Роль для человечества и окружающей среды
Крайне трудно недооценить значение зеленых водорослей в биосфере, ведь поглощая выделяемый животными и людьми углекислый газ, они отдают кислород, без которого жизнь на планете становится невозможной по определению. Благодаря этой уникальной особенности, растения широко используются не только на земле, но и в космических полетах, а также на подводных суднах. Кроме того, их часто применяют в качестве своеобразного индикатора уровня загрязнения воды, воздуха и других важнейших экосистем.
Огромное количество всевозможных витаминов, минералов и микроэлементов, которые входят в состав этих растительных организмов, позволяет активно использовать определенные их виды в кулинарии (особенно любят этот компонент жители стран Юго-Восточной Азии), а также в косметологии. Самые эффективные маски, крема, сыворотки и глины для обертывания, как правило, обязательно содержат экстракты этих удивительных растений.
Кроме того, морские водоросли играют огромную роль в фармакологии, ведь их полезные качества и свойства активно используются для изготовления самых разных лекарственных препаратов для внутреннего и наружного применения.
Зеленые водоросли, подготовка к ЕГЭ по биологии
Самый обширный отдел водорослей, включающий от 13 000 до 20 000 видов. Обитают в основном в пресных водоемах, имеют зеленую окраску вследствие преобладания хлорофилла a и b по количеству над другими пигментами (каротиноидами, ксантофиллами). Этот отдел включает в себя одноклеточные, многоклеточные и колониальные формы. Большинство из них растет на глубине 20-40 метров.
Клеточная стенка зеленых водорослей образована целлюлозой, запасное питательное вещество - крахмал. У многих представителей в жизненном цикле наблюдается чередование полового поколения (гаметофита) и бесполого (спорофита).
Хламидомонада
Хламидомонада - одноклеточная двужгутиковая зеленая водоросль, обитающая в лужах, пресных водоемах, прудах. Форма клетки грушевидная. На переднем конце тела имеет два жгутика, за счет которых активно движется.
Светочувствительный глазок (стигма) помогает хламидомонаде занять наиболее освещенное место для активного процесса фотосинтеза, который идет в хроматофоре. Сократительные (пульсирующие) вакуоли клетки удаляют избыток постоянно поступающей внутрь воды, таким образом, они поддерживают осмотическое давление на уровне, необходимом для жизни.
Хламидомонада имеет чашевидный хроматофор с пиреноидом - округлой белковой гранулой, содержащей фермент, который участвует в синтезе сахаров. Вокруг пиреноида запасается крахмал.
Размножение хламидомонады
Размножается преимущественно бесполым путем. Половой процесс - конъюгация.
- Бесполое размножение
- Половое размножение
При благоприятных условиях (летом) размножается бесполым путем с помощью зооспор. Хламидомонада (n) дважды делится митотически без разрыва материнской оболочки, в результате образуются 4 клетки (n). Они растут, у каждой из них развивается жгутик, появляется глазок и клеточная стенка. С течением времени материнская оболочка, окружающая клетки, разрывается, и зооспоры выходят во внешнюю среду. Из каждой зооспоры развивается взрослая клетка.
Рассмотрим изогамный половой процесс, при котором гаметы не отличаются по строению, внешнему виду, одинаково подвижны.
Половое размножение активируется при наступлении неблагоприятных условий (пересыхание водоема, понижение температуры внешней среды). Внутри хламидомонады (n) путем митоза образуются половые клетки - гаметы (n). Запомните, что в половом размножении всегда участвуют половые клетки ;)
Гаметы (n) разных хламидомонад попарно сливаются, в результате чего образуется зигота (2n), которая покрывается плотной защитной оболочкой - цистой. При благоприятных условиях зигота (2n) делится мейозом, по итогам которого образуются 4 хламидомонады (n).
Красный снег
Красный снег - явление, характерное для приполярных областей Земли, также встречается на высоких горах. Снег приобретает нехарактерную красную окраску, связанную с массовым размножением Хламидомонады снежной, клетки которой содержат красный каротиноид - астаксантин. Для особей этого вида благоприятными являются низкие температуры, при температуре выше +4 °С они погибают.
Хлорелла
Хлорелла - одноклеточная зеленая водоросль без жгутиков, обитающая в самых разных средах: на сырой почве, на стволах деревьев, скалах, в соленой и пресной воде. Ее скопления хорошо заметны в виде налета зеленого цвета.
Клетка содержит чашевидный хроматофор (имеет вид сильно вырезанной чаши), запасающий крахмал. Хлорелла отличается быстрым темпом деления клеток, в связи с этим ее используют для получения кормов. Фотосинтез у нее также идет очень интенсивно. Эта водоросль одной из первых побывала в космосе, ее используют на космических кораблях для получения кислорода.
Размножение осуществляется только бесполым путем, содержимое материнской клетки делится митотически на 4 или 8 дочерних клеток, после чего оболочка материнской клетки рвется, и дочерние клетки выходят наружу, развиваются во взрослых особей, после чего снова делятся.
Спирогира
Спирогира - многоклеточная нитчатая зеленая водоросль. Скопления нитей спирогиры на поверхности рек и прудов образуют тину.
Хроматофор у спирогиры спиралевидный, представлен в виде одной или нескольких лент, опоясывающих клетку в пристенном слое цитоплазмы. В клетке содержится крупное ядро, расположенное в центре и подвешенное на тяжах цитоплазмы.
Размножается бесполым и половым путями.
- Бесполое
- Половое
Бесполое (вегетативное) размножение может осуществляться частями таллома: нить водоросли разрывается на отдельные участки, или даже клетки, которые дают начало новому организму.
Половой процесс - конъюгация. Две нити водоросли располагаются параллельно, клетки сближаются, у них образуются боковые выросты. При соприкосновении боковых выростов между клетками разных нитей водорослей образуется копуляционный канал, по которому происходит перемещение содержимого одной клетки (n) в другую (n), после чего сливаются цитоплазмы и ядра, образуя зигоспору (2n).
После периода покоя зигоспора (2n) делится мейозом, образуются четыре клетки (n), из которых только одна прорастает в новую особь, а три остальных - погибают.
Кладофора
Кладофора - многоклеточная нитчатая зеленая водоросль. Ее ветвящиеся нити непрочно прикреплены к субстрату, от которого часто отрываются. Хроматофор имеет вид сеточки (сетчатый). Бесполое размножение осуществляется с помощью зооспор, половое размножение в форме изогамии.
Улотрикс
Улотрикс - многоклеточная нитчатая зеленая водоросль. Обитает в пресной и морской воде, образует на подводных объектах зеленый налет - тину. Хроматофор в виде незамкнутого кольца (пояска), содержит пиреноид. Преимущественно размножается бесполым путем, с помощью четырехжгутиковых зооспор. Есть возможность полового размножения по типу изогамии.
В цикле развития улотрикса преобладает гаметофит (n) - вегетативное гаплоидное поколение. Также заметьте, что гаметы улотрикса (n) образуются из клеток слоевища (n) путем митоза.
Плеврококк
Скорее всего, любой гетеротроф сделает ошибку, первый раз встретив это название :) Уж слишком сильно оно смахивает на название бактерий, таких как стафилококки, стрептококки. Запомните и не ошибайтесь: плеврококк - зеленая водоросль. Плеврококк имеет клетки шаровидной формы, они могут быть одиночные или соединенные в группы. Видимые вакуоли в клетке отсутствуют, хроматофор в виде пластинки, не содержит пиреноидов.
Плеврококк распространен повсеместно, способен вынести полное пересыхание. Образует зеленый налет на стволах деревьев, поверхности скал и почве.
Вольвокс
«Вольвокс» означает «катящийся». Представляет собой зеленую подвижную колониальную водоросль, имеющую шаровидную форму. Одна колония вольвокса может достигать 3мм, а по количеству клеток - 200 до 10 тысяч.
Клетки расположены на периферии, соединены между собой тяжами цитоплазмы - протоплазматическими нитями, обеспечивают движение колонии и питание. В центре колонии имеется полость, занятая слизью. Каждая из клеток на периферии имеет два жгутика, обращенных во внешнюю среду, клетки напоминают хламидомонаду.
Вольвокс играет очень важное эволюционное значение, и помогает сделать вывод о том, что развитие живых организмов от одноклеточных форм к многоклеточным происходило через колониальные формы.
Большая часть клеток в колонии вольвокса вегетативные. Вегетативное размножение вольвокса происходит с помощью дочерних колоний внутри материнской, особыми клетками - партеногонидиями. Эти клетки делятся митозом перпендикулярно поверхности шара. В результате образуется пластинка, которая выворачивается и образует дочерний шар. Дочерние шары разрастаются, при этом происходит разрыв материнского организма (шара).
Половой процесс происходит в специализированных местах - антеридиях, где развиваются сперматозоиды (n), и оогониях, где созревают яйцеклетки (n). Сперматозоид проникает в оогоний, образуется зигота, или ооспора (2n). При благоприятных условиях зигота делится мейозом, образуются клетки вольвокса (n), которые затем делятся множеством митотических делений.
Таким образом, основная форма существования клеток в колонии вольвокса - гаплоидна (n), диплоидна в жизненном цикле только зигота (2n).
Сине-зеленые водоросли
Спешу предупредить об очень частом заблуждении! Сине-зеленые водоросли - это вовсе не водоросли, их по-другому называют цианобактерии. Они представляют собой отдел крупных грамотрицательных бактерий, которые способны выделять кислород в процессе фотосинтеза.
Эволюционно сине-зеленые водоросли - очень древние микроорганизмы, которые возникли в архее. Им отведена крайне важна роль: они являются первыми фотосинтезирующими организмами. Благодаря им 2 млрд. лет назад в атмосфере Земли впервые появился кислород.
У них отсутствуют жгутики, они могут иметь нитчатую или колониальную форму, или же быть одноклеточными. Относительно крупные размеры цианобактерий и сходство в строении с водорослями было изначальной причиной их рассмотрения в составе растений. На настоящее время доказано сходство цианобактерий с остальными бактериями.
© Беллевич Юрий Сергеевич 2018-2020
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Зеленые водоросли: особенности строения, представители вида и их количество, основное предназначение
Зеленые водоросли представляют собой обширнейшую группу живых микроорганизмов, ткани которых практически не подлежат какой-либо дифференциации, поэтому их по-прежнему относят к группе низших растений, включающих в себя, помимо обозначенного вида, бактерии, лишайники и грибы.
Зеленые водоросли считаются низшими организмами на ряду с бактериями и грибамиБиологический вид Chlorophyta
Отдел Chlorophyta включает в себя огромный список зеленых водорослей, который по предварительным оценкам ученых-исследователей исчисляется тысячами видов растений (приблизительное их число варьируется в крайне широком диапазоне от 13 до 20 тысяч).
Именно поэтому определить точное количество их видов на сегодняшний день невозможно и главная причина такого положения заключается в сложности дифференциации этих растительных организмов.
Представители отдела
Практически все виды таллома можно встретить среди зеленых водорослей. Представителями этого отдела являются не только простейшие одноклеточные и многоклеточные колониальные водоросли планктонного типа, неспособные сопротивляться течению водных масс, но и одно- и многоклеточный фитобентос, отличительная черта которого заключается в его среде обитания — океанском, морском или речном грунте.
Не все представители класса зеленых водорослей имеют зеленый цветЕдинственным исключением являются одно- и многоклеточные растения ризоподиального типа с их сложной структурой строения и отличным талломом.
Тем не менее именно рассматриваемый отдел официально признан самым обширным во всем растительном мире водоемов. Несмотря на название, далеко не все представители обладают слоевищем (телом) зеленого оттенка. К примеру, такой представитель вида, как трентеполия, имеет клетки оранжево-красного пигмента, обусловленные накоплением каротина и всевозможных производных этого полезного вещества.
Встречаются среди этих водорослей и растения нитчатого типа, главное отличие которых сводится не к преобладающему пигменту другого цвета, а к среде обитания. Если в начале своего жизненного пути их можно отнести к бентосам, так как нитчатые водоросли имеют свойство крепиться к водоемному грунту, то со временем они пускаются в свободное плавание, фактически становясь планктонными водорослями, формирующими многочисленные шары и маты из своих колоний.
Классы зеленых водорослей
Из-за сложности идентификации талломов растительных организмов, входящих в отдел водорослей зеленого цвета, а также их более чем внушительного количества, ученым до сих пор так и не удалось создать какой-то единый их реестр или подробный классификатор. Тем не менее в настоящее время выделяют пять основных классов зеленых водорослей, к которым относят:
- ульвофициевые;
- брипсодовые;
- хлорофициевые;
- требуксиевые;
- празиновые.
В класс Ulvophyceae входит не менее тысячи видов растений, преимущественно обитающих в морской среде. Примечательно, что к этому классу относят не только нитчатые и пластинчатые водоросли, но и некоторые виды лишайников. Что же касается морских его представителей, которых абсолютное большинство, то характерной их особенностью является наличие известковых микрочастиц. В свою очередь, этот класс делится на два порядка:
Класс Bryopsidophyceae насчитывает около пятисот видов растительных организмов, обладающих талломом неклеточного типа. Все эти водоросли относятся к единому порядку с одноименным названием Бриопсидовые, преобладающему в пресных водах. В свою очередь, этот порядок делится на три основных рода: бриопсис, кодиум и каулерпа, отличающихся друг от друга не только строением, но и средой обитания.
Класс одноклеточных водорослей Chlorophyceae насчитывает около 2500 видов, которые относятся к единому порядку вольвоксовых. При этом он делится на два рода: хламидомонада (не менее 500 видов) и вольвокс (около 2 тысяч). Первый род, который также является бесполым, имеет свойство размножаться посредством деления клетки. В то же время вольвокс, который по праву считается наиболее высокоорганизованным представителем этой группы, способен организовывать огромные колонии, насчитывающие тысячи живых организмов.
Хламидомонада относится к классу зеленых водорослейЕще один класс Trebouxiophyceae состоит из 170 видов одноклеточных организмов, имеющих преимущественно коккоидную форму, которые могут обитать как на земляном грунте, так и на дне пресных озер и рек.
Этот класс классифицируется по двум порядкам на хлорелловые и требуксиевые водоросли, и если первый тип представляет собой неподвижные шарики, для которых естественной средой обитания являются водоемы, то второй входит в состав лишайников, встречающихся на древесной коре либо ведущих свободный образ жизни.
Название последнего класса Prasinophyceae переводится с древнегреческого языка как «зеленый» и на этот раз речь также идет об одноклеточных организмах с коккоидным, жгутиковым или пальмеллоидным типом строения. Делится этот класс на два порядка: пирамимонадовые, которые нередко провоцируют цветение пресных и соленых вод, и хлородендровые подвижного и неподвижного типа со слизкими ножками для крепления к твердым поверхностям. Водоросли последнего порядка часто имеют тот самый красный пигмент из-за наличия каротина. Кроме того, они могут паразитировать, всю жизнь обитая в теле морских червей.
Структура, циклы и предназначение
Несмотря на огромное количество видов зеленых водорослей, которые поражают своим разнообразием, ученые смогли выделить несколько общих характеристик этих живых растительных организмов.
Особенности строения и жизненный цикл
Клетки пластид, которые входят в состав водорослей, чаще всего имеют чашевидную форму, и, помимо классического зеленого, они включают в свой состав желтый (лютеин) и красно-оранжевые пигменты каротиновой группы (зеаксантин, виолаксантин, неоксантин и пр.). Сам хлоропласт имеет особое внутреннее включение — пиреноид, который пронизан 2−6 ограниченными мембранами компартментами, именуемые тилакоидами.
Жизненный цыкл зеленых водорослей зависит от среды их обитанияПри этом зеленые водорослевые пластиды имеют сдвоенные мембраны, что свойственно красным водорослям и другим более развитым растительным организмам.
Клетки большинства видов рассматриваемой группы имеют сократительные вакуоли, под которыми подразумеваются одномембранные органоиды с клеточным соком в средней части. Не исключено и наличие стигмы — внутриклеточного органоида с ярко-красными пятнами, благодаря которому организм способен двигаться по направлению к или от светового источника.
Практически идентичные по своей структуре жгутики нередко имеют разную длину, при том что их может быть два и более. Эти изоконты не имеют боковых волосков (мастигонем), однако, это вовсе не означает, что у водорослевых жгутиков полностью отсутствуют любые волоски или чешуйки.
Фактором усиленного роста водорослей является повышение температуры среды их обитанияЖизненный цикл этих растительных организмов напрямую зависит от их вида и среды обитания. Всего же ученые выделяют три основных цикла их развития:
- Гаплобионтный (зиготическая редукция), для которого свойственно высвобождение гаметы через пору материнской клетки и ее дальнейшее слияние с другой гаметой через трубку. Далее происходит преобразование в зигоспору, в которой после периода покоя образуется 4 зооспоры с последующим формированием полиэдра в каждой из них.
- Гапло-диплобионтный (спорическая редукция), при котором вышедшие из материнской клетки гаметы сливаются в воде, образуя не двух, как в гаплобиотном цикле, а четырехжгутиковые зиготы, свободно парящие в водных массах. Следующий этап предполагает прикрепление зиготы к любой твердой поверхности и ее покрытие плотным слоем, после чего происходит формирование кодиолума, уходящего на длительный покой. Повторная активация этих дубинкообразных клеток, как правило, становится возможной при наступлении благоприятных условий. В итоге из кодиолума выделяется от 4 до 16 спор, формирующих новые нити и все повторяется вновь.
- Диплобионтный (гаметическая редукция), характеризующийся делением крупного ядра, которое образуется при оседании планозиготы и формировании отдельного нитчатого таллома.
Наиболее благоприятным фактором для развития водорослей является повышение температуры, поэтому совсем неудивительно, что их бурный рост можно наблюдать в теплых водах и при наступлении весеннего и летнего сезонов.
Роль для человечества и окружающей среды
Крайне трудно недооценить значение зеленых водорослей в биосфере, ведь поглощая выделяемый животными и людьми углекислый газ, они отдают кислород, без которого жизнь на планете становится невозможной по определению. Благодаря этой уникальной особенности, растения широко используются не только на земле, но и в космических полетах, а также на подводных суднах. Кроме того, их часто применяют в качестве своеобразного индикатора уровня загрязнения воды, воздуха и других важнейших экосистем.
Огромное количество всевозможных витаминов, минералов и микроэлементов, которые входят в состав этих растительных организмов, позволяет активно использовать определенные их виды в кулинарии (особенно любят этот компонент жители стран Юго-Восточной Азии), а также в косметологии. Самые эффективные маски, крема, сыворотки и глины для обертывания, как правило, обязательно содержат экстракты этих удивительных растений.
Кроме того, морские водоросли играют огромную роль в фармакологии, ведь их полезные качества и свойства активно используются для изготовления самых разных лекарственных препаратов для внутреннего и наружного применения.
ПредыдущаяАквариумНитчатые зеленые водоросли в аквариуме: способы борьбы с нитевидными растениями, как избавиться от нитчатки
СледующаяАквариумХламидомонада: где обитает зеленая водоросль и чем питается, способ размножения
Отличная статья 0
Строение водорослей, подготовка к ЕГЭ по биологии
Водоросли относятся к низшим растениям, наиболее примитивным: у них отсутствует разделение организма на стебель, корень и листья. Спешу заметить, что термин "низшие растения" - отжившее понятие, использовавшееся в ботанике до второй половины XX века.
Современная биология не считает дифференциацию тканей определяющим различием, сейчас существенным считают фундаментальные различия в строение клеток, обмене веществ. Тем не менее, во многих устаревших пособиях этот термин используется, и я обязан предупредить вас о нем.
Наука о водорослях называется альгология (от лат. alga — морская трава, водоросль и греч. λόγος — учение).
Среди водорослей есть одноклеточные и многоклеточные, некоторые водоросли достигают в длину 100-200 метров. Способ питания водорослей автотрофный: они синтезируют органические вещества в процессе фотосинтеза. Солнечный свет, проходя через толщу воды, рассеивается, что делает фотосинтез с увеличением глубины все труднее и труднее. Поэтому кроме хлорофилла они часто имеют и другие пигменты.
Клетки водорослей характеризуются наличием клеточной стенки (из целлюлозы и гликопротеинов - от греч. glykys сладкий (углеводы) + греч. prōtos — первый, важнейший (белок)) Органоиды располагаются в цитоплазме (син. - внеядерной протоплазме), где также располагается(-ются) один или несколько хроматофоров. Размножение происходит бесполым, вегетативным или половым путем.
Тело водорослей представлено слоевищем (син. - талломом) - недифференцированным скоплением клеток. С помощью ризоидов (от др.-греч. ῥίζα — корень и εἶδος — вид) водоросли прикрепляются к субстрату (камням, коралловым полипам), функцию всасывания ризоиды не выполняют. У водорослей отсутствуют настоящие ткани, механических тканей нет, так как таллом водоросли поддерживается (парит) в толще воды. Нет проводящих тканей: каждая клетка имеет доступ к воде напрямую, так что в клетку из окружающей воды поступает кислород, а в воду удаляется углекислый газ.
Хроматофор (от греч. chroma - цвет и phoros - несущий) - органелла в клетке водоросли, аналогичная хлоропласту и осуществляющая фотосинтез. Отличается от хлоропласта упрощенным строением, меньшим размером и иным составом хлорофилла. Внешне отличаются между собой по форме, хроматофор может быть: чашевидный, спиралевидный, в виде незамкнутых колец, цилиндрические, лентовидные, дисковидные. В хроматофорах находятся пигменты, которые придают окраску растению.
Система вакуолей в клетках водорослей развита отлично, в подвижных клетках водорослей можно обнаружить пульсирующие (сократительные) вакуоли. Их основная функция - поддержание постоянного осмотического давления внутри клетки. Вообразите: в глубине океана находится клетка водоросли, в которую постоянно поступает много воды. Если бы не было таких сократительных вакуолей, то клетка просто лопнула бы, но их работа обеспечивает удаление избытка воды.
Также у многих подвижных водорослей в клетках присутствует светочувствительный глазок (стигма), что обуславливает их чувствительность к свету - фототаксис. Подвижные водоросли стремятся занять как можно более освещенное место, чтобы активно шел процесс фотосинтеза.
Жизненный цикл водорослей
Жизненные циклы водорослей разнообразны, обусловлены рядом экологических факторов. Мы разберем жизненный цикл на примере зеленой водоросли ульвы (морского салата).
Для начала отметим, что в целом жизненный цикл водорослей представляет собой чередование двух фаз: гаплоидной (гаметофита) и диплоидной (спорофита). Гаплоидной фазой называется фаза, при которой клеточные ядра содержат непарный (половинный) набор хромосом. К гаплоидной фазе всегда принадлежат гаметы: сперматозоиды, спермии (отличающиеся от сперматозоидов отсутствием жгутика), яйцеклетки.
При слиянии двух гамет: яйцеклетки (n) и спермия (n) образуется зигота (2n) из которой развивается спорофит (2n), таким образом, в спорофите восстанавливается диплоидный набор хромосом. В зооспорангии на спорофите в результате мейоза образуются зооспоры (n), которые делятся митозом, порастают и образуют мужские и женские гаметофиты (n). Клетки гаметофитов делятся митозом, образуются гаметы (n), которые сливаются в зиготу (2n), цикл замыкается.
Типы половых процессов
У водорослей выделяют несколько типов полового процесса:
- Изогамия - копулирующие элементы (гаметы) не отличаются друг от друга, подвижны
- Анизогамия - от греч. anisos неравный и gamos брак (гетерогамия) - при таком типе копулирующие элементы различаются по размерам, форме, величине, поведению
- Оогамия - от др. греч. ᾠόν яйцо и γάμος брак - копулирующие элементы резко отличаются друг от друга: крупная женская гамета без жгутиков обычно с мужской мелкой подвижной гаметой. Допустимо считать оогамию в некотором смысле подтипом анизогамии.
Особо стоит выделить тип полового процесса - конъюгацию. Конъюгация отличается тем, что сливаются не гаметы, а обычные вегетативные клетки, лишенные жгутиков. Клетки соединяются друг с другом с помощью боковых выростов, формируется копуляционный (конъюгационный) канал, по которому содержимое из одной клетки перетекает в другую - образуется зигоспора. В дальнейшем из зигоспоры развивается новая водоросль.
Отметим, что зооспора представляет собой подвижную клетку, которая способна двигаться в воде с помощью жгутиков. Образуется она в зооспорангии. Зооспора участвует в бесполом размножении у многих водорослей и простейших грибов. У некоторых водорослей имеются апланоспоры (гр. aplanes неподвижный + spora семя) - неподвижные безжгутиковые споры. Зооспоры и апланоспоры выходят в окружающую среду, разрывая стенки спорангия, в котором они находятся.
Значение водорослей
В Мировом океане водоросли составляют основную часть биомассы. Именно они являются главными продуцентами (производителями) органического вещества, преобразуя в ходе фотосинтеза энергию солнечного света в энергию химических связей. Значение водорослей для человека трудно переоценить: содержащиеся в них вещества необходимы для нормального роста и развития животных и человека (к примеру, морская капуста (ламинария) отличается большим содержанием йода.)
© Беллевич Юрий Сергеевич 2018-2020
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
общая информация и характеристики :: SYL.ru
Зеленые водоросли – один из самых многочисленных отделов водорослей. Они широко распространены на нашей планете и включают до 20 000 видов. Прежде всего их объединяет цвет основного пигмента, в остальном же они очень разные. Давайте узнаем, какие водоросли – зеленые. Какие виды и классы к ним относятся? Какими чертами они обладают?
Главные признаки водорослей
Водорослями мы обычно называем все растения, живущие в воде. Влагу они действительно любят, однако это далеко не единственный их признак. Они представляют собой обширную группу разных по происхождению организмов, которые объединены некоторыми общими особенностями:
- наличие хлорофилла;
- фотоавтотрофность;
- отсутствие разделения тела на функциональные части;
- отсутствие покровной оболочки;
- отсутствие отчетливой проводящей системы.
Водоросли обитают во влажной местности. Они могут жить в почве, на поверхности земли, на коре растений, в морях, океанах, болотах и других водоемах. Они являются важными участниками экологических и биологических процессов. Именно с водорослей начинаются морские и некоторые наземные пищевые цепочки.
По типу питания они являются фотоавтотрофами. Это значит, что они умеют производить органические вещества из неорганических и делают это, используя энергию света и углекислый газ. Своей деятельностью они производят примерно половину всего объема кислорода, который производится растениями.
Их тело называется таллом, или слоевищем. В отличие от тела высших растений оно не разделяется на разные органы и ткани. И хотя внешне водоросли могут иметь видимые стебли, корни, листья - это всего лишь обман. Все их части состоят из клеток одного типа, которые выполняют одни и те же функции.
Какие бывают водоросли?
Известно больше 100 тысяч видов водорослей. Они подразделяются на отделы: бурые, диатомовые, красные, зеленые водоросли. Все они являются эукариотами, так как их клетки содержат ядро. Однако науке известны и сине-зеленые водоросли, которые являются прокариотическими организмами. За способность к фотосинтезу их когда-то причисляли к растениям, но теперь они считаются бактериями и встречаются под названиями «цианей» или «цианобактерий».
Зеленые водоросли относятся к царству растений и включают организмы самых разнообразных форм и размеров. Чаще всего они обитают в пресных водоемах и увлажненных регионах, но встречаются и в соленых, и в солоноватых водах.
Существует несколько классов зеленых водорослей:
- ульфоциевые;
- бриопсидовые;
- хлорофициевые;
- требуксиевые;
- празиновые;
Их размеры колеблются от нескольких микрометров до двух метров. Характерный зеленый цвет придает им высокое содержание хлорофилла. Кроме него, водоросли содержат пигмент лютеин, неоксантин. Из-за присутствия каротиноидов некоторые из них имеют красноватый или оранжевый оттенок.
Внешний вид и строение
Внутреннее и внешнее строение зеленых водорослей очень разнообразно. Они могут быть одноядерными и многоядерными, состоять из разного количества клеток или вообще иметь неклеточный таллом. Некоторые из них не прикрепляются к поверхностям и свободно живут в водной среде. Другие же прочно закреплены на предметах и различных субстратах.
Среди зеленых водорослей есть такие, которые состоят всего из одной клетки. Разглядеть их без микроскопа не получится, ведь их размер чрезвычайно мал. Зато летом и весной можно наблюдать, как от их активного размножения вода в лужах, прудах и болотах окрашивается в зеленый цвет.
Многоклеточные зеленые водоросли больше напоминают обыкновенные высшие растения. Их тело может состоять из множества нитей, образовывать подобие стебельков и листочков. Промежуточный вариант между многоклеточными и одноклеточными представляют колониальные. Они представляют собой группу соединенных между собой клеток или организмов. Несмотря на объединение, они могут сохранять свою самостоятельность и по-разному реагировать на раздражители. В случае разрыва колонии они спокойно существуют дальше и способны образовывать новые группы при помощи деления своих клеток.
Отношения с другими организмами
Зеленые водоросли могут существовать практически во всех влажных местах. Они встречаются в почве, на тенистой стороне камней и даже в снегах высоких гор и арктического пояса Земли. Если стакан с водой оставить на несколько дней в солнечном месте, то они появятся и там.
Водоросли легко вступают в симбиотические отношения с другими организмами. Они прекрасно сосуществуют с моллюсками, губками, простейшими, гидрами и плоскими червями. С грибами их сотрудничество оказалось настолько продуктивным, что даже получило отдельное название – лишайник. Когда-то такое объединение считали целостным растительным организмом, например мхом. В образовании лишайников участвует около 80 водорослей, из которых чаще всего встречаются представители рода требуксия.
Одноклеточные зеленые водоросли порой развиваются в шерсти или кожном покрове млекопитающих, а также на высших растениях. Их присутствие не всегда полезно для других. Порой они просто паразитируют и приводят к заболеваниям хозяина. Они способны вызвать мастит у рогатого скота, становятся причиной некоторых кожных заболеваний человека, истощают растения чая, приводя к пожелтению листьев.
Хламидомонада
Род хламидомонада объединяет больше 500 видов зеленых водорослей, обитающих преимущественно в пресных водоемах. Это одноклеточные организмы грушевидной или овальной формы. Они оснащены специальными светочувствительными глазками и парой жгутиков, вращательные движения которых помогают хламидомонадам перемещаться в воде к более освещенным местам.
Большинство из них обитают в небольших, хорошо прогретых водоемах и могут способствовать их цветению. Самый необычный представитель – хламидомонада снежная, живущая при низких температурах. Она обитает в снегах и льдах, и благодаря пигменту астаксантину окрашивает их в розоватый оттенок.
Спирогира
Спирогира является самой распространенной многоклеточной водорослью. Она окрашена в ярко-зеленый цвет и состоит из множества тонких ниточек различной длины. Спирогира встречается как в пресных, так и в соленых водах. Она появляется в медленно текущих и застойных водоемах, в аквариумах, за которыми неправильно ухаживают. Вместе с другими водорослями она образует липкую тину, наощупь напоминающую вату.
Вольвокс
Вольвокс – подвижные колониальные водоросли, обитающие в стоячих пресных водоемах. В период массового размножения они способствуют цветению воды, окрашивая ее в зеленый цвет. Известно около 20 видов вольвокса.
Колония вольвокса выглядит как зеленый шар размером максимум 3 мм. В каждый из таких шаров входит от 10 до 200 тысяч микроскопических клеток, которые соединяются нитками из протоплазмы. Как и у хламидомонады, у них имеются жгутики для передвижения внутри водоема. Клетки не одинаковы и разделяются по своей специфике. Одни являются вегетативными, другие генеративными и принимают участие в половом размножении.
Ульва
Род ульва представляет морские зеленые водоросли, обитающие в пределах субтропического и умеренного поясов земного шара. Они являются многоклеточными организмами с ветвистым пластинчатым талломом размером от 30 сантиметров до 1,5 метра. У основания тела водоросли находятся нитевидные образования, ризоиды, при помощи которых они цепляются за различные поверхности.
Ульва нуждается в солнечном свете, поэтому обитает на небольших глубинах. Она служит кормом морских животных и является отличным местом для откладывания икры рыбами. Некоторые виды ульвы люди употребляют в пищу. В кулинарии она больше известна как морской салат.
Нителла
Водоросли рода нителла широко распространены в пределах Северного полушария. Они обитают в пресных водоемах Азии, Европы и Северной Америки. Внешне водоросли сложно отличить от высших растений. Их таллом состоит из тоненьких стебельков с небольшими узлами, от которых по кругу отходит 5-7 узких и тонких листочков.
Они любят хорошо освещенные места и теплую температуру воды - в пределах 20-28 градусов. Растение довольно неприхотливо и способно делать воду прозрачнее, отчего его часто заводят в аквариумах. В естественных водоемах разросшаяся нителла становится убежищем небольших рыбешек и местом для их нереста.
Каулерпа
Бриопсидовые водоросли рода каулерпа обитают преимущественно в теплых тропических и субтропических морях. Внешне они очень разнообразны и визуально делятся на листья, стебли и корни. Они могут быть ветвистыми или напоминать по виду грибы. Несмотря на видимую сложность, их внутреннее строение очень простое. Таллом водорослей состоит всего из одной клетки, которая содержит множество ядер. У некоторых видов она может сильно разрастаться, достигая 2-3 метра в размере.
Каулерпа неподвижна и всегда прикрепляется к какой-нибудь поверхности. Часто она селится на дне водоемов, цепляясь к илу или песку. Она также растет на подводных камнях и коралловых рифах. Благодаря своим размерам, водоросли этого рода являются крупнейшими одноклеточными организмами в мире. Из-за необычного вида они пользуются популярностью, поэтому их часто выращивают в аквариумах. В странах Юго-Восточной и Восточной Азии некоторые виды каулерп употребляют в пищу.
список основных видов и описание
Зеленые водоросли представляют собой обширнейшую группу живых микроорганизмов, ткани которых практически не подлежат какой-либо дифференциации, поэтому их по-прежнему относят к группе низших растений, включающих в себя, помимо обозначенного вида, бактерии, лишайники и грибы.
Зеленые водоросли считаются низшими организмами на ряду с бактериями и грибамиБиологический вид Chlorophyta
Отдел Chlorophyta включает в себя огромный список зеленых водорослей, который по предварительным оценкам ученых-исследователей исчисляется тысячами видов растений (приблизительное их число варьируется в крайне широком диапазоне от 13 до 20 тысяч).
Именно поэтому определить точное количество их видов на сегодняшний день невозможно и главная причина такого положения заключается в сложности дифференциации этих растительных организмов.
Представители отдела
Практически все виды таллома можно встретить среди зеленых водорослей. Представителями этого отдела являются не только простейшие одноклеточные и многоклеточные колониальные водоросли планктонного типа, неспособные сопротивляться течению водных масс, но и одно- и многоклеточный фитобентос, отличительная черта которого заключается в его среде обитания — океанском, морском или речном грунте.
Не все представители класса зеленых водорослей имеют зеленый цветЕдинственным исключением являются одно- и многоклеточные растения ризоподиального типа с их сложной структурой строения и отличным талломом.
Тем не менее именно рассматриваемый отдел официально признан самым обширным во всем растительном мире водоемов. Несмотря на название, далеко не все представители обладают слоевищем (телом) зеленого оттенка. К примеру, такой представитель вида, как трентеполия, имеет клетки оранжево-красного пигмента, обусловленные накоплением каротина и всевозможных производных этого полезного вещества.
Если Вам понравилось видео — поделись с друзьями:
Поделиться на Facebook
Поделиться в ВК
Поделиться в ОК
Поделиться в Twitter
Встречаются среди этих водорослей и растения нитчатого типа, главное отличие которых сводится не к преобладающему пигменту другого цвета, а к среде обитания. Если в начале своего жизненного пути их можно отнести к бентосам, так как нитчатые водоросли имеют свойство крепиться к водоемному грунту, то со временем они пускаются в свободное плавание, фактически становясь планктонными водорослями, формирующими многочисленные шары и маты из своих колоний.
Классы зеленых водорослей
Из-за сложности идентификации талломов растительных организмов, входящих в отдел водорослей зеленого цвета, а также их более чем внушительного количества, ученым до сих пор так и не удалось создать какой-то единый их реестр или подробный классификатор. Тем не менее в настоящее время выделяют пять основных классов зеленых водорослей, к которым относят:
- ульвофициевые;
- брипсодовые;
- хлорофициевые;
- требуксиевые;
- празиновые.
В класс Ulvophyceae входит не менее тысячи видов растений, преимущественно обитающих в морской среде. Примечательно, что к этому классу относят не только нитчатые и пластинчатые водоросли, но и некоторые виды лишайников. Что же касается морских его представителей, которых абсолютное большинство, то характерной их особенностью является наличие известковых микрочастиц. В свою очередь, этот класс делится на два порядка:
- Улотриксовые, чаще всего обитающие в водоемах с пресной водой, прикрепляясь к каким-либо предметам и образуя многочисленные колонии в виде низкорослых кустиков. Бесполые растения этого порядка имеют свойство размножаться двумя способами, первый из которых подразумевает деление одной нити на несколько, в то время как для второго свойственно образование зооспор в материнских клетках с дальнейшим их выбросом и распространением.
- Ульвовые, которые встречаются во всех морях мира, предпочитая в первую очередь соленые водоемы с теплой водой. Отличительной характеристикой этих водорослей с талломом ярко-зеленого цвета является наличие ножки для крепления к морскому грунту, а также разнообразие строения (слоевище растений может иметь как трубчатую, так и мешковидную или пластинчатую форму).
Если Вам понравилось видео — поделись с друзьями:
Поделиться на Facebook
Поделиться в ВК
Поделиться в ОК
Поделиться в Twitter
Класс Bryopsidophyceae насчитывает около пятисот видов растительных организмов, обладающих талломом неклеточного типа. Все эти водоросли относятся к единому порядку с одноименным названием Бриопсидовые, преобладающему в пресных водах. В свою очередь, этот порядок делится на три основных рода: бриопсис, кодиум и каулерпа, отличающихся друг от друга не только строением, но и средой обитания.
Класс одноклеточных водорослей Chlorophyceae насчитывает около 2500 видов, которые относятся к единому порядку вольвоксовых. При этом он делится на два рода: хламидомонада (не менее 500 видов) и вольвокс (около 2 тысяч). Первый род, который также является бесполым, имеет свойство размножаться посредством деления клетки. В то же время вольвокс, который по праву считается наиболее высокоорганизованным представителем этой группы, способен организовывать огромные колонии, насчитывающие тысячи живых организмов.
Хламидомонада относится к классу зеленых водорослейЕще один класс Trebouxiophyceae состоит из 170 видов одноклеточных организмов, имеющих преимущественно коккоидную форму, которые могут обитать как на земляном грунте, так и на дне пресных озер и рек.
Этот класс классифицируется по двум порядкам на хлорелловые и требуксиевые водоросли, и если первый тип представляет собой неподвижные шарики, для которых естественной средой обитания являются водоемы, то второй входит в состав лишайников, встречающихся на древесной коре либо ведущих свободный образ жизни.
Название последнего класса Prasinophyceae переводится с древнегреческого языка как «зеленый» и на этот раз речь также идет об одноклеточных организмах с коккоидным, жгутиковым или пальмеллоидным типом строения. Делится этот класс на два порядка: пирамимонадовые, которые нередко провоцируют цветение пресных и соленых вод, и хлородендровые подвижного и неподвижного типа со слизкими ножками для крепления к твердым поверхностям. Водоросли последнего порядка часто имеют тот самый красный пигмент из-за наличия каротина. Кроме того, они могут паразитировать, всю жизнь обитая в теле морских червей.
Если Вам понравилось видео — поделись с друзьями:
Поделиться на Facebook
Поделиться в ВК
Поделиться в ОК
Поделиться в Twitter
Структура, циклы и предназначение
Несмотря на огромное количество видов зеленых водорослей, которые поражают своим разнообразием, ученые смогли выделить несколько общих характеристик этих живых растительных организмов.
Особенности строения и жизненный цикл
Клетки пластид, которые входят в состав водорослей, чаще всего имеют чашевидную форму, и, помимо классического зеленого, они включают в свой состав желтый (лютеин) и красно-оранжевые пигменты каротиновой группы (зеаксантин, виолаксантин, неоксантин и пр.). Сам хлоропласт имеет особое внутреннее включение — пиреноид, который пронизан 2−6 ограниченными мембранами компартментами, именуемые тилакоидами.
Жизненный цыкл зеленых водорослей зависит от среды их обитанияПри этом зеленые водорослевые пластиды имеют сдвоенные мембраны, что свойственно красным водорослям и другим более развитым растительным организмам.
Клетки большинства видов рассматриваемой группы имеют сократительные вакуоли, под которыми подразумеваются одномембранные органоиды с клеточным соком в средней части. Не исключено и наличие стигмы — внутриклеточного органоида с ярко-красными пятнами, благодаря которому организм способен двигаться по направлению к или от светового источника.
Практически идентичные по своей структуре жгутики нередко имеют разную длину, при том что их может быть два и более. Эти изоконты не имеют боковых волосков (мастигонем), однако, это вовсе не означает, что у водорослевых жгутиков полностью отсутствуют любые волоски или чешуйки.
Фактором усиленного роста водорослей является повышение температуры среды их обитанияЖизненный цикл этих растительных организмов напрямую зависит от их вида и среды обитания. Всего же ученые выделяют три основных цикла их развития:
- Гаплобионтный (зиготическая редукция), для которого свойственно высвобождение гаметы через пору материнской клетки и ее дальнейшее слияние с другой гаметой через трубку. Далее происходит преобразование в зигоспору, в которой после периода покоя образуется 4 зооспоры с последующим формированием полиэдра в каждой из них.
- Гапло-диплобионтный (спорическая редукция), при котором вышедшие из материнской клетки гаметы сливаются в воде, образуя не двух, как в гаплобиотном цикле, а четырехжгутиковые зиготы, свободно парящие в водных массах. Следующий этап предполагает прикрепление зиготы к любой твердой поверхности и ее покрытие плотным слоем, после чего происходит формирование кодиолума, уходящего на длительный покой. Повторная активация этих дубинкообразных клеток, как правило, становится возможной при наступлении благоприятных условий. В итоге из кодиолума выделяется от 4 до 16 спор, формирующих новые нити и все повторяется вновь.
- Диплобионтный (гаметическая редукция), характеризующийся делением крупного ядра, которое образуется при оседании планозиготы и формировании отдельного нитчатого таллома.
Наиболее благоприятным фактором для развития водорослей является повышение температуры, поэтому совсем неудивительно, что их бурный рост можно наблюдать в теплых водах и при наступлении весеннего и летнего сезонов.
Если Вам понравилось видео — поделись с друзьями:
Поделиться на Facebook
Поделиться в ВК
Поделиться в ОК
Поделиться в Twitter
Роль для человечества и окружающей среды
Крайне трудно недооценить значение зеленых водорослей в биосфере, ведь поглощая выделяемый животными и людьми углекислый газ, они отдают кислород, без которого жизнь на планете становится невозможной по определению. Благодаря этой уникальной особенности, растения широко используются не только на земле, но и в космических полетах, а также на подводных суднах. Кроме того, их часто применяют в качестве своеобразного индикатора уровня загрязнения воды, воздуха и других важнейших экосистем.
Огромное количество всевозможных витаминов, минералов и микроэлементов, которые входят в состав этих растительных организмов, позволяет активно использовать определенные их виды в кулинарии (особенно любят этот компонент жители стран Юго-Восточной Азии), а также в косметологии. Самые эффективные маски, крема, сыворотки и глины для обертывания, как правило, обязательно содержат экстракты этих удивительных растений.
Кроме того, морские водоросли играют огромную роль в фармакологии, ведь их полезные качества и свойства активно используются для изготовления самых разных лекарственных препаратов для внутреннего и наружного применения.
Если Вам понравилось видео — поделись с друзьями:
Поделиться на Facebook
Поделиться в ВК
Поделиться в ОК
Поделиться в Twitter
(Visited 1 201 times, 1 visits today)
Вконтакте
Google+
Знакомство с зелеными водорослями
Знакомство с зелеными водорослями
«Зеленые водоросли» - самая разнообразная группа водорослей, насчитывающая более 7000 виды, растущие в самых разных средах обитания. «Зеленые водоросли» - это парафилетическая группа , поскольку исключает Plantae. Как растения, зеленые водоросли содержат две формы хлорофилла, которые они используют для улавливания световой энергии для производства сахаров, но в отличие от растения они прежде всего водные.Потому что они водные и производятся их собственная пища, эти организмы называются « водорослей », наряду с некоторые члены Хромиста, Родофита и фотосинтезирующие бактерии, даже при том, что они не имеют близких отношений ни с одной из этих групп.
На изображении выше показан густой рост морского салата ( Ulva ), растет в водоеме в порту Беркли. Это морской вид «зеленые водоросли» часто обнаруживаются прикрепленными к камням и обнаженными во время отлива.
Посетите Галерею доисторических водорослей, чтобы увидеть морские растения, которые жили между 700 и 400 миллионами лет назад. .
Нажмите на кнопки ниже, чтобы узнать больше о «Зеленых водорослях».
Изображение выше любезно предоставлено Википедией, используется с CC BY-SA 3.0
Посетите Висконсин, чтобы увидеть фотографии различных зеленых водорослей или попробуйте Protist База данных изображений для информации о Хламидомонада, Тетрасельмис, Halosphaera и Пирамимоны - все «зеленые водоросли».Дополнительные изображения зеленого водоросли доступны от Tavole di Botanica sistematica.
Duke Univeristy содержит хламидомонады Центр генетики, включающий базы данных и учебные материалы.
.
Зеленых водорослей - Простая английская Википедия, бесплатная энциклопедия
Зеленые водоросли - микроскопические протисты. Их можно найти в любой природной воде: соленой, пресной и солоноватой. Группа в настоящее время используется в классификации реже. Хотя большинство из них произошли от общего предка, некоторые - нет. Некоторые из их потомков, наземные растения не входят в группу, поэтому группа не является монофилетической.
Зеленые водоросли - это большая группа водорослей, от которых произошли высшие растения. [1]
Зеленые водоросли включают одноклеточные и колониальные жгутиконосцы, обычно, но не всегда с двумя жгутиками на клетку, а также различные колониальные, кокковидные и нитчатые формы. У харалов, ближайших родственников высших растений, происходит полная дифференциация тканей. Есть около 6000 видов зеленых водорослей. [2] Многие виды живут большую часть своей жизни одноклеточными, другие виды образуют колонии или длинные волокна.
.Производство водорода. Зеленые водоросли как источник энергии
- Copyright © 2001 Американское общество физиологов растений
Реферат
Газообразный водород считается идеальным топливом для мира, в котором удалось снизить уровень загрязнения воздуха, остановить глобальное потепление и обеспечить экономическую защиту окружающей среды. Водород и электричество могут объединиться, чтобы предоставить привлекательные возможности для транспорта и производства электроэнергии. Взаимное преобразование между этими двумя формами энергии предполагает использование водорода на месте для выработки электроэнергии, при этом электрическая сеть служит для транспортировки энергии, использования распределения и регенерации водорода по мере необходимости.Сложной проблемой при внедрении H 2 в качестве источника энергии для будущего является возобновляемое и экологически безопасное производство большого количества газа H 2 . Таким образом, процессы, которые в настоящее время носят концептуальный характер или находятся на стадии разработки в лаборатории, необходимо поощрять, проверять на осуществимость и иным образом применять для коммерциализации.
С момента пионерского открытия Гаффроном и его коллегами более 60 лет назад (Gaffron, 1939; Gaffron and Rubin, 1942) способность одноклеточных зеленых водорослей производить газ H 2 при освещении была в основном биологической диковинкой.Исторически активность выделения водорода у зеленых водорослей индуцировалась при предварительной анаэробной инкубации клеток в темноте (Greenbaum, 1982; Roessler, Lien, 1984; Happe and Naber, 1993; Schulz, 1996). Фермент гидрогеназа экспрессируется при такой инкубации и катализируется с высокой удельной активностью опосредованной светом эволюции H 2 . Сообщается, что мономерная форма фермента относится к классу гидрогеназ Fe (Voordouw et al., 1989; Adams, 1990; Meyer and Gagnon, 1991; Happe et al., 1994), кодируется в ядре одноклеточных зеленых водорослей. Однако зрелый белок локализован и функционирует в строме хлоропластов (Happe et al., 1994). Поглощение света фотосинтетическим аппаратом необходимо для образования газообразного водорода, поскольку энергия света способствует окислению молекул воды, высвобождению электронов и протонов и эндергоническому переносу этих электронов к ферредоксину. Фотосинтетический ферредоксин (PetF) служит физиологическим донором электронов для Fe-гидрогеназы и, таким образом, связывает Fe-гидрогеназу с цепью переноса электронов в хлоропласте зеленых водорослей (Florin et al., 2001).
В этих условиях активность гидрогеназы носит временный характер (она длится от нескольких секунд до нескольких минут), потому что, помимо электронов и протонов, светозависимое окисление воды влечет за собой высвобождение молекулярного O 2 . Кислород является мощным ингибитором гидрогеназы Fe (Ghirardi et al., 2000). Текущие технологические разработки в этой области еще не преуспели в преодолении этой взаимоисключающей природы реакций фоторождения O 2 и H 2 .Таким образом, физиологическое значение и роль Fe-гидрогеназы в зеленых водорослях, которые обычно растут в условиях аэробного фотосинтеза, долгое время оставались загадкой. Учитывая чувствительность Fe-гидрогеназы к O 2 и преобладающие окислительные условия окружающей среды на Земле, были заданы вопросы о том, является ли гидрогеназа чем-то большим, чем пережитком эволюционного прошлого хлоропласта зеленых водорослей, и может ли этот фермент и процесс фотосинтеза может быть использован для получения газа H 2 в коммерческих целях (Zhang et al., 2001). Тем не менее, способность зеленых водорослей к фотосинтетическому образованию газа H 2 привлекла внимание научного сообщества из-за фундаментальной и практической важности этого процесса. Ниже приводится подробный список свойств и перспектив фотосинтеза в производстве зеленых водорослей H 2 , а также проблем, с которыми сталкиваются современные технологии:
(a) Фотосинтез в зеленых водорослях может происходить с эффективностью преобразования фотонов ≥80 % (Лей и Маузералл, 1982).
(b) Микроводоросли могут производить H 2 фотосинтетически, с эффективностью преобразования фотонов ≥80% (Greenbaum, 1988).
(c) Молекулярный O 2 действует как мощный и эффективный переключатель, с помощью которого отключается производственная деятельность H 2 .
(d) Эту несовместимость при одновременном фотопроизводстве O 2 и H 2 не удалось преодолеть за 60 лет соответствующих исследований.
Помимо описанной выше фотосистемы II (ФСII) -зависимой H 2 фотоэволюция, которая включает воду в качестве источника электронов и производит 2: 1 стехиометрические количества H 2 : O 2 , альтернативный вариант. Механизм описан в литературе (Gfeller and Gibbs, 1984).После анаэробной инкубации водорослей в темноте и последующей индукции гидрогеназы электроны для фотосинтетического аппарата образуются в результате катаболизма эндогенного субстрата и сопутствующего окислительного метаболизма углерода в зеленых водорослях. Электроны от такого эндогенного катаболизма субстратов поступают в фотосинтетическую цепь переноса электронов между двумя фотосистемами и, вероятно, на уровне пула пластохинонов. Поглощение света PSI и последующий перенос электронов повышает окислительно-восстановительный потенциал этих электронов до окислительно-восстановительного эквивалента ферредоксина и гидрогеназы, что позволяет генерировать молекулярный H 2 (Гиббс и др., 1986). В присутствии ингибитора PSII 3- (3,4-дихлорфенил) -1,1-диметилмочевины (DCMU) в этом процессе образуются стехиометрические количества H 2 : CO 2 2: 1. Таким образом, после достаточно продолжительной анаэробной инкубации культуры в темноте первоначально высокие скорости продукции H 2 могут быть обнаружены при освещении водорослей в присутствии DCMU, ингибитора PSII (Happe and Naber, 1993; Florin et al. , 2001).
ДВУХЭТАПНЫЙ ФОТОСИНТЕЗ И H 2 ПРОИЗВОДСТВО В ЗЕЛЕНЫХ ВОДОРОСЛЯХ
Недавняя работа показала, что недостаток серы в среде роста Chlamydomonas reinhardtii вызывает специфическое, но обратимое снижение скорости кислородного фотосинтеза (Wykoff et al., 1998), но не влияет на скорость митохондриального дыхания (Melis et al., 2000). В герметизированных культурах дисбаланс во взаимосвязи фотосинтеза-дыхания из-за депривации S привел к чистому потреблению кислорода клетками, вызывающим анаэробиоз в среде для выращивания, условие, которое автоматически вызвало продукцию H 2 водорослями (Melis et al., 2000 ). В ходе этой недавней работы было показано, что экспрессия гидрогеназы Fe может быть индуцирована на свету, пока анаэробиоз сохраняется в культуре (Ghirardi et al., 2000; Melis et al., 2000). В таких условиях можно было получать и накапливать значительные объемы газа H 2 с использованием зеленой водоросли C. reinhardtii в устойчивом процессе, который можно было использовать непрерывно в течение нескольких дней. Таким образом, прогресс был достигнут за счет обхода чувствительности гидрогеназы Fe к O 2 за счет временного разделения реакций фотообразования O 2 и H 2 , то есть за счет так называемого «двухстадийного фотосинтеза и H 2 production »(Melis et al., 2000). Новое применение этого двухэтапного протокола выявило наличие ранее неизвестных метаболических, регуляторных и электронных путей у зеленой водоросли C. reinhardtii (Zhang et al., 2001), что привело к значительному и устойчивому влиянию света. выброс газа H 2 клетками. На рисунке 1 показана фотография изолированной и продуцирующей H 2 культуры C. reinhardtii , лишенной S, на которой выделяются выделяющиеся пузырьки H 2 , которые ненадолго задерживаются на стенках и шейке Roux. бутылка.
 Рис.1.
Рис.1. Водород-продуцирующая культура C. reinhardtii . Пузырьки водорода выходят к поверхности жидкой среды. Газ сливается через шприц (вставленный в середину силиконовой пробки) и через тефлоновую трубку собирается в перевернутую бюретку и измеряется методом вытеснения воды. Фотография любезно предоставлена Майклом Барнсом (Офис президента Калифорнийского университета, Окленд).
Этот метод может служить инструментом для выяснения метаболизма зеленых водорослей, связанного с водородом.При дальнейшей доработке метод может также использоваться для получения газа H 2 для топливной и химической промышленности. Временная последовательность событий в этом двухэтапном фотосинтезе и процессе производства H 2 представлена ниже:
(а) Зеленые водоросли выращивают фотосинтетически на свету (нормальный фотосинтез) до тех пор, пока они не достигнут плотности от 3 до 6 миллионов клеток на мл -1 в культуре.
(b) Депривация серы навязывается клеткам в среде для выращивания либо путем тщательного ограничения поступления серы в среду, чтобы она была полностью израсходована, либо путем разрешения клеткам концентрироваться в камере для выращивания перед заменой среды на среду, в которой отсутствует серные питательные вещества.Клетки отвечают на эту депривацию S, фундаментально изменяя фотосинтез и клеточный метаболизм, чтобы выжить (Davies et al., 1996; Hell, 1997; Zhang et al., 2001).
(c) Депривация S оказывает совершенно иное влияние на клеточную активность фотосинтеза и дыхания. Активность оксигенного фотосинтеза снижается квазиэкспоненциально с периодом полувыведения от 15 до 20 часов до значения менее 10% от его первоначальной скорости (Wykoff et al., 1998). Однако способность клеточного дыхания остается довольно постоянной в течение периода депривации S (Melis et al., 2000). Как следствие, абсолютная активность фотосинтеза ниже уровня дыхания примерно через 24 часа S-депривации. Следуя этой точке пересечения между фотосинтезом и дыханием, запечатанные культуры C. reinhardtii , лишенные S, быстро потребляют весь растворенный кислород и становятся анаэробными (Ghirardi et al., 2000), даже если они находятся под постоянным освещением.
(d) В условиях S-депривации запечатанные (анаэробные) культуры C.reinhardtii выделяет газ H 2 на свету, но не в темноте. Объем и скорость продукции фотосинтетического H 2 контролировали по накопившемуся газу H 2 в перевернутой бюретке, измеряя по объему вытеснения воды. Скорость от 2,0 до 2,5 мл, продукция H 2 L -1 культура h -1 поддерживалась в период от 24 до 70 часов. После этого ставка постепенно снижалась.
(e) В ходе такого образования газа H 2 клетки потребляли значительные количества внутреннего крахмала и белка (Zhang et al., 2001). Такие катаболические реакции, по-видимому, прямо или косвенно поддерживают процесс производства H 2 .
(f) Анализ профиля выбранных фотосинтетических белков показал резкое снижение количества Rubisco в зависимости от времени в S-депривации, более постепенное снижение уровня белков PSII и PSI, а также изменение состава световых лучей. уборочный комплекс.
(e) Микроскопические наблюдения показали отчетливые морфологические изменения у C.reinhardtii во время S-депривации и производства H 2 . Клетки эллипсоидной формы (нормальный фотосинтез) уступили место более крупным и сферическим клеткам на начальных (0–24 ч) стадиях S-депривации и продукции H 2 с последующим уменьшением клеточной массы при более длительных (24–120 ч) S депривации и H 2 раз производства (Zhang et al., 2001).
Краткое изложение текущего состояния дел в этой области приведено ниже:
(а) Отсутствие серы в среде роста водорослей действует как метаболический переключатель, который избирательно и обратимо выключает фотосинтетическое производство O 2 .
(b) В присутствии S зеленые водоросли осуществляют нормальный фотосинтез (окисление воды, выделение O 2 и накопление биомассы). В отсутствие S и O 2 фотосинтез у C. reinhardtii переходит в продуктивный режим H 2 .
(c) Обратимое применение переключателя (присутствие / отсутствие S) позволяет водорослям чередовать производство O 2 и производство H 2 (чередование стадий; Ghirardi et al., 2000), таким образом обходя несовместимость и взаимоисключающую природу реакций образования O 2 и H 2 .
(d) Взаимодействие между кислородным фотосинтезом, митохондриальным дыханием, катаболизмом эндогенного субстрата и переносом электронов через гидрогеназный путь является важным для этого опосредованного светом процесса производства H 2 .
(e) Высвобождение газа H 2 служит для поддержания базовых уровней активности переноса электронов хлоропластов и митохондрий для выработки АТФ, который необходим для выживания организма в условиях длительного стресса, вызванного депривацией серы (см. Ниже) .
ФИЗИОЛОГИЯ H 2 ПРОИЗВОДСТВО В ЗЕЛЕНЫХ ВОДОРОСЛЯХ
Исторически Ганс Гаффрон сделал первое наблюдение за водородным метаболизмом в зеленых водорослях (Gaffron, 1939, 1944). При воздействии водорода на анаэробно адаптированные клетки он наблюдал поглощение водорослями молекулярного H 2 и сопутствующее снижение CO 2 в темноте. Обратная реакция, например о производстве водорода на свету впервые сообщалось у зеленой водоросли Scenedesmus obliquus (Gaffron and Rubin, 1942).Высокие скорости эволюции H 2 можно было измерить на свету за короткие промежутки времени (от нескольких секунд до нескольких минут). Электроны генерировались либо при фотохимическом окислении воды ФСII, что приводит к одновременному образованию O 2 и H 2 (Spruit, 1958; Greenbaum et al., 1983), либо при окислении эндогенного субстрата ( Рис.2), подающие электроны в тилакоидную мембрану с одновременным высвобождением в среду CO 2 (Kessler, 1974; Bamberger et al., 1982). Известно, что C. reinhardtii может фотообразовать водород, когда ФСII блокируется DCMU, но после добавления 2,5-дибром-3-метил-6-изопропил-пара-бензохинона выделение H 2 не происходит (Stuart и Gaffron, 1972), который блокирует функцию комплекса цитохрома bf . В анаэробных условиях в присутствии DCMU накопленные восстанавливающие эквиваленты в результате ферментативного катаболизма водорослей не могут быть окислены посредством дыхания, поскольку конечный акцептор электронов O 2 отсутствует.Белковый комплекс НАД (Ф) Н-редуктазы, который поставляет электроны в пул пластохинонов, недавно был обнаружен во многих хлоропластах сосудистых растений (Shinozaki et al., 1986; Kubicki et al., 1996; Sazanov et al., 1998), но пока только из зеленой водоросли Nephroselmis olivacea (Turmel et al., 1999). Тем не менее, эксперименты с ингибиторами дали доказательства в поддержку локализованной на тилакоидной мембране НАД (Ф) Н-редуктазы у C. reinhardtii (Godde and Trebst, 1980), предполагая, что электроны, полученные при окислении эндогенного субстрата, могут поступать в пластохинон бассейн (рис.2). После этого при поглощении света PSI электроны направляются к ферредоксину. Последний является эффективным донором электронов для гидрогеназы Fe, которая эффективно объединяет эти электроны с протонами с образованием молекулярного H 2 (Florin et al., 2001).
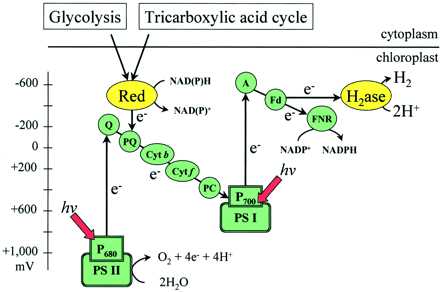 Рис.2.
Рис.2. Связанные с гидрогеназой пути переноса электронов в зеленых водорослях. Электроны могут возникать либо на ФСII при фотоокислении воды, либо в пуле пластохинонов при окислении эндогенного субстрата клетки (например,г. через гликолиз и цикл трикарбоновых кислот). Электроны в цепи переноса электронов транспортируются через PSI к ферредоксину, который служит физиологическим донором электронов для гидрогеназы Fe. Р 680 , Реакционный центр ФСII; П 700 , реакционный центр ПСИ; Q - первичный акцептор электронов ФС II; A - первичный акцептор электронов PSI; PQ, пластохинон; Цит, цитохром; ПК, пластоцианин; Fd, ферредоксин; Красный - НАД (Ф) Н оксидоредуктаза; H 2 аза, гидрогеназа; ФНР, ферредоксин-НАДФ + редуктаза.
Физиология продукции H 2 при депривации S имеет много общего и некоторых отличий от процесса, описанного выше. Лишенные серы и запечатанные культуры C. reinhardtii становятся анаэробными на свету из-за значительного и специфического замедления активности ФСII, развивающего O 2 , за которым следует автоматическая индукция гидрогеназы Fe и фотосинтез H 2 производство. Биохимический анализ показал, что одновременно с производственным процессом H 2 содержание крахмала и белка в клетках постепенно снижалось (Zhang et al., 2001). Такой катаболический путь (пути) может приводить к образованию восстановителя, который подает электроны на тилакоидную мембрану, возможно, через хлоропластный NAD (P) H-зависимый процесс (Gfeller and Gibbs, 1984). Что еще более важно, катаболизм крахмала должен также создавать субстрат для митохондриального дыхания клетки. Митохондриальное дыхание поглощает небольшие количества O 2 , которые выделяются из-за остаточной активности фотосинтеза, и, таким образом, обеспечивает поддержание анаэробиоза в культуре (рис.3). Таким образом, физиология продукции H 2 за счет S-депривации включает скоординированное взаимодействие между:
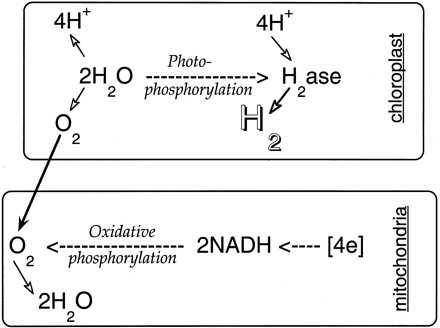 Рис.3.
Рис.3. Скоординированный фотосинтетический и респираторный транспорт электронов и сопряженное фосфорилирование во время продукции H 2 . Фотосинтетический перенос электронов доставляет электроны при фотоокислении воды гидрогеназе, что приводит к фотофосфорилированию и образованию H 2 . Кислород, образующийся в результате этого процесса, служит для управления окислительным фосфорилированием во время митохондриального дыхания.Электроны для последнего ([4e]) образуются при катаболизме эндогенного субстрата, который дает восстановитель и CO 2 . Высвобождение молекулярного H 2 хлоропластом делает возможным устойчивое действие этой скоординированной функции фотосинтеза-дыхания у зеленых водорослей и позволяет непрерывно генерировать АТФ двумя биоэнергетическими органеллами в клетке.
(a) Кислородный фотосинтез, то есть остаточная активность ФСII по генерации электронов при окислении воды.Эти электроны переносятся через фотосинтетическую цепь переноса электронов и, в конечном итоге, поступают в гидрогеназу Fe, тем самым внося свой вклад в продукцию H 2 .
(b) Митохондриальное дыхание поглощает весь кислород, генерируемый остаточным фотосинтезом, и, таким образом, поддерживает анаэробиоз в культуре.
(c) Катаболизм эндогенного субстрата, включая крахмал, белок и, вероятно, липидный катаболизм, дает субстрат, пригодный для работы окислительного фосфорилирования в митохондриях и, возможно, для NAD (P) H-зависимого транспорта электронов в хлоропласте, причем оба эти фактора способствуют выработке столь необходимого АТФ.
(d) Транспорт электронов по гидрогеназному пути и последующее высвобождение газа H 2 водорослями поддерживает базовый уровень фотосинтеза и, следовательно, респираторный транспорт электронов для генерации АТФ (рис. 3) и, таким образом, обеспечивает выживание организма в условиях длительного стресса.
Очевидно, что необходимы дополнительные исследования, чтобы проанализировать четырехстороннее взаимодействие и сложные взаимосвязи между процессами кислородного фотосинтеза, митохондриального дыхания, катаболизма эндогенного субстрата и транспорта электронов через гидрогеназный путь, ведущий к продукции H 2 .Тем не менее, открытие устойчивого производства H 2 , которое обходит чувствительность обратимой гидрогеназы к O 2 , является значительным достижением в этой области. Это может привести к использованию зеленых водорослей для производства газа H 2 в качестве чистого и возобновляемого топлива. Однако фактическая скорость накопления газа H 2 составляла в лучшем случае от 15% до 20% фотосинтетической способности клеток, когда последняя основана на способности выделять O 2 в физиологических условиях (Melis et al., 2000). Относительно низкая скорость производства H 2 предполагает, что есть возможности для значительного улучшения выхода процесса на порядок величины. Точно так же необходимо внести другие улучшения для оптимизации процесса в условиях массового культивирования водорослей. Например, необходимо решить оптические проблемы, связанные с размером антенны хлорофилла и кривой светонасыщения фотосинтеза (Melis et al., 1999), прежде чем зеленые водоросли смогут достичь высокой эффективности фотосинтетического преобразования солнечной энергии в массовой культуре.Более того, необходимо обеспечить непрерывность процесса, поскольку производство H 2 за счет S-лишения водорослей не может длиться вечно. Урожайность начинает стабилизироваться примерно через 70 часов S-депривации. Примерно через 100 часов лишения серы водоросли должны вернуться к нормальному фотосинтезу, чтобы омолодиться за счет восполнения эндогенного субстрата (Ghirardi et al., 2000).
НОВЫЙ ТИП Fe-ГИДРОГЕНАЗЫ В ЗЕЛЕНЫХ ВОДОРОСЛЯХ
Первоначально гидрогеназы Feбыли клонированы из анаэробных микроорганизмов и простейших, продуцирующих H 2 (Meyer and Gagnon, 1991; Bui and Johnson, 1996; Ахманова и др., 1998; Vignais et al., 2001). Эти ферменты позволяют поддерживать ферментативный метаболизм в анаэробных условиях за счет использования протонов (вместо O 2 ) в качестве концевого акцептора электронов и поддерживать процесс, выделяя газ H 2 (Peters, 1999). Гидрогеназы Fe отличаются своей чувствительностью к CO и высокой ферментативной активностью, которая в 100 раз выше, чем у гидрогеназ NiFe. Структура гидрогеназ Fe из Clostridium pasteurianum и Desulfovibrio desulfuricans была недавно выяснена с помощью рентгеновской кристаллографии (Peters et al., 1998; Николет и др., 1999). Эти белки имеют многодоменную структуру с многочисленными кластерами [Fe-S], включая новый тип кластера [Fe-S] (H-кластер) в каталитическом сайте фермента. H-кластер состоит из обычного комплекса [4Fe-4S], связанного атомом серы остатка Cys с уникальным биядерным субкластером железо-сера (Adams and Stiefel, 2000). Считается, что высококонсервативные аминокислотные остатки, содержащие четыре лиганда Cys и несколько гидрофобных аминокислотных остатков в активном центре, участвуют в образовании каналов H + и H 2 , тем самым соединяя каталитический сайт (расположенный глубоко внутри белка матрикс) на поверхность белка (Vignais et al., 2001).
Несмотря на открытие водородного метаболизма у зеленых водорослей более 60 лет назад и большой интерес к биологической эволюции H 2 с тех пор, попытки клонировать и охарактеризовать ген гидрогеназы этих фотосинтезирующих организмов оказались безуспешными. Однако недавно гены гидрогеназы были выделены и описаны в литературе из зеленых водорослей S. obliquus (Florin et al., 2001), C. reinhardtii (Happe and Kaminski, 2001) и Chlorella fusca . (Т.Хаппе, личное сообщение). Показано, что все три гена принадлежат к классу гидрогеназ Fe. Однако они показали новые структурные свойства и предположили уникальную биохимическую функцию. Интересно, что гены гидрогеназы Fe не могут быть обнаружены у цианобактерий, свободноживущих предков пластид, что повышает вероятность нецианобактериального происхождения гидрогеназ водорослей.
Гидрогеназы Fe из зеленых водорослей представляют собой мономерные белки массой примерно от 45 до 50 кДа, очищенные до гомогенности (Roessler and Lien, 1984; Happe and Naber, 1993).Кодируемые ядром полипептиды синтезируются в цитозоле как белки-предшественники, но зрелый белок локализован в строме хлоропласта (Happe et al., 1994). Транзитный пептидный домен, который направляет Fe гидрогеназы из цитоплазмы через оболочку хлоропласта в строму хлоропласта, был идентифицирован в N-концевой области фермента (Florin et al., 2001). Нацеливающий на хлоропласт домен белка, вероятно, расщепляется локализованной в строме пептидазой в консервативном сайте расщепления.Никакие дополнительные гены, которые могли бы участвовать в биосинтезе и / или сборке гидрогеназ Fe, еще не идентифицированы ни у зеленых водорослей, ни у других микроорганизмов, содержащих гидрогеназы Fe.
Генетические данные по Fe гидрогеназам зеленых водорослей (HydA) выявляют уникальные особенности этого класса ферментов (Florin et al., 2001). Они представляют собой самые маленькие из известных белков Fe-гидрогеназы со значительно укороченным N-концевым доменом и консервативным C-концевым доменом, который содержит каталитический сайт.Функционально важный С-конец последовательности HydA очень похож на таковой у других гидрогеназ Fe анаэробных микроорганизмов. Четыре высококонсервативных остатка Cys координируют специальный кластер [6Fe-6S] (H-кластер) в каталитическом сайте (рис. 4). Ряд дополнительных аминокислотных остатков определяет среду активного сайта. Было высказано предположение, что 12 в основном гидрофобных аминокислотных остатков могут играть роль в защите H-кластера от окружающей водной среды (Peters et al., 1998). Десять остатков строго консервативны, тогда как два остатка варьируются в пределах семейства гидрогеназ Fe (Ser-232 и Ile-268 в C. pasteurianum ; Ala-109 и Thr-145 в D. desulfuricans ; Ala-38 и Thr- 74 в C. reinhardtii ; и Ala-44 и Thr-80 в S. obliquus ). Однако последовательности зеленых водорослей включают вставку из 16-45 аминокислот, которая отсутствует в бактериальных последовательностях и которая образует внешнюю пептидную петлю в полностью собранном белке.Эта дополнительная пептидная петля в гидрогеназе зеленой водоросли может участвовать в электростатическом связывании природного донора электронов ферредоксина.
 Рис.4.
Рис.4. Сравнительная схема полипептидов гидрогеназы Fe из выбранных организмов, продуцирующих H 2 . Зрелый полипептид гидрогеназы Fe изображен зеленым линейным прямоугольником. Вертикальные стрелки и соответствующие параллелограммы внутри линейного прямоугольника показывают положение консервативных аминокислот в протеине гидрогеназы.Остатки Cys, которые играют роль в координации кластеров [Fe-S], показаны желтым. Консервативные аминокислоты, не содержащие Cys, показаны синим цветом. Вставка 16 и 45 уникальных аминокислотных остатков в гидрогеназу Fe S. obliquus и C. reinhardtii , соответственно, показана светло-зеленым цветом.
В N-концевом домене бактериальных и других неводорослевых Fe-гидрогеназ обнаружено, что ряд остатков Cys, которые явно отсутствуют у аналогов зеленых водорослей, связывают дополнительные железо-серные кластеры.Во всех негидрогеназах Fe ферредоксин-подобный домен (F-кластер) координирует два кластера [4Fe-4S] (Peters et al., 1998; Adams and Stiefel, 2000). Дополнительные кластеры железной серы были обнаружены в Fe-гидрогеназах C. pasteurianum (рис. 4), Thermotoga maritima (Verhagen et al., 1999) и Nyctotherus ovalis (Ахманова и др., 1998). F-кластер в этих организмах отвечает за перенос электронов от донора электронов (в основном ферредоксина) к H-кластеру (Nicolet et al., 2000). Эти дополнительные центры [Fe-S] отсутствуют в гидрогеназах водорослей, что указывает на новый путь переноса электронов от донорного ферредоксина PetF к кластеру гидрогеназы H. Отсутствие таких дополнительных центров [Fe-S] и соответственно более короткий полипептид Fe-гидрогеназы зеленых водорослей значительно сокращает расстояние от сайта донорства электронов ферредоксина до H кластера (Florin et al., 2001). В этом отношении внешняя пептидная петля гидрогеназ водорослей может компенсировать недостающие домены.Положительно заряженные аминокислоты в петлевой структуре могут служить доменом для связывания ферредоксина. Таким образом, это может помочь ориентировать отрицательно заряженный ферредоксин для облегчения связывания и эффективного переноса электронов между ферредоксином и гидрогеназой. Такое взаимодействие является предпосылкой значимого связывания фермента с цепью переноса электронов в хлоропластах.
РЕГУЛЯЦИЯ БИОСИНТЕЗА ГИДРОГЕНАЗ В ЗЕЛЕНЫХ ВОДОРОСЛЯХ
С момента открытия водородного метаболизма у зеленых водорослей (Gaffron, 1939) стало известно, что активность гидрогеназы может быть обнаружена только при анаэробной инкубации клеток.Недавняя работа (Happe et al., 1994; Happe and Kaminski, 2001) предоставила молекулярное понимание этого свойства и показала, что гидрогеназа Fe быстро индуцируется на уровне транскрипции при анаэробиозе. Интересно, что это явление индукции было использовано в качестве руководящего принципа для клонирования гена гидрогеназы Fe в зеленых водорослях (Happe and Kaminski, 2001). Был использован специальный генетический метод, так называемая супрессивная субтрактивная гибридизация, основанная на сравнении популяций мРНК из клеток, выращенных в аэробных условиях и инкубированных в анаэробных условиях.Этот подход позволил выделить ряд клонов кДНК, которые экспрессировались только в анаэробных условиях. Один из этих клонов кодировал гидрогеназу Fe C. reinhardtii (Happe, Kaminski, 2001).
Эукариотические зеленые водоросли, выращенные в фотоавтотрофных условиях окружающей среды, не потребляют и не производят молекулярный водород, что свидетельствует об отсутствии экспрессии гена гидрогеназы Fe. Вестерн-блоттингом (Happe et al., 1994) и нозерн-блоттингом (Happe and Kaminski, 2001) было показано, что ни белковые, ни генные транскрипты не могут быть обнаружены в фотоавтотрофных (аэробных) условиях окружающей среды.Однако экспрессия гена гидрогеназы Fe индуцируется при инкубации клеток в анаэробных условиях в темноте, например барботирование аргоном (Happe and Naber, 1993) или при инкубации в условиях депривации S, например условия, которые избирательно снижают активность PSII и O 2 evolution (Melis et al., 2000; Zhang et al., 2001). Экспрессия гена гидрогеназы Fe начинается в течение 10 мин после того, как среда становится анаэробной и, вероятно, регулируется на уровне транскрипции.Эти результаты предполагают, что кислород прямо или косвенно действует как положительный супрессор экспрессии гена гидрогеназы Fe на уровне транскрипции. В отсутствие кислорода подавление экспрессии гена гидрогеназы облегчается и беспрепятственно протекает в клетках. Такой запуск, очевидно, не зависит от метода, используемого для установления анаэробиоза, а также от освещения и фотосинтеза в зеленых водорослях, пока анаэробиоз сохраняется внутри клетки.
СКОЛЬКО H 2 МОЖНО ЖДАТЬ ОТ МАССОВОЙ КУЛЬТУРЫ ЗЕЛЕНЫХ ВОДОРОСЛЕЙ?
Применение двухэтапного фотосинтеза и производственного протокола H 2 к массовой культуре зеленых водорослей может обеспечить коммерчески жизнеспособный метод получения возобновляемого водорода. В таблице I представлены предварительные оценки максимально возможного урожая H 2 зелеными водорослями на основе яркости солнца и характеристик фотосинтеза зеленых водорослей. Расчеты основывались на интегральной светимости Солнца в безоблачный весенний день.В средних широтах весной это повлечет за собой доставку примерно 50 моль фотонов m −2 d −1 (таблица I, строка 1). Принято считать, что перенос электронов двумя фотосистемами и по гидрогеназному пути для производства 1 моль H 2 требует поглощения и использования минимум 5 моль фотонов в фотосинтетическом аппарате (таблица I, строка 2). На основе этих «оптимальных» допущений можно рассчитать, что зеленые водоросли могут производить максимум 10 моль (20 г) H 2 на м 2 посевных площадей в день.Если бы урожай такой величины можно было достичь в массовой культуре, это стало бы жизнеспособным и прибыльным методом производства возобновляемой продукции H 2 .
Таблица I.Выход фоторождения водорода зелеными водорослями
Однако этот оптимистичный сценарий не может быть реализован с использованием современных ноу-хау. На эту технологию производства H 2 напрямую влияют три биологически «серых зоны». (а) Выход продукции H 2 , достигаемый в настоящее время в лаборатории, соответствует только 15-20% измеренной способности фотосинтетического аппарата к переносу электронов (Melis et al., 2000). (b) Оптические свойства поглощения света зелеными водорослями накладывают ограничение с точки зрения эффективности преобразования солнечного излучения в хлоропласте водорослей. Это связано с тем, что зеленые водоросли дикого типа оснащены большой антенной из хлорофилла, собирающей свет, чтобы поглощать как можно больше солнечного света. Под прямыми и яркими солнечными лучами они могут тратить до 60% поглощенного излучения (Neidhardt et al., 1998; Melis et al., 1999). Эта эволюционная черта может быть полезной для выживания организма в дикой природе, где свет часто ограничивает, но не способствует фотосинтетической продуктивности массовой культуры зеленых водорослей.Это оптическое свойство клеток может еще больше снизить производительность коммерческой фермы по производству H 2 . (c) Текущая необходимость в цикле культивирования между двумя стадиями (нормальный фотосинтез в присутствии S, чередующийся с продукцией H 2 при депривации S) вводит «время простоя» в отношении продукции H 2 . «Время простоя» неизбежно приведет к дальнейшему снижению производительности производственного процесса H 2 . Таким образом, согласно существующим технологиям, предполагается, что фактический выход продукции H 2 будет ниже, чем у теоретического максимума, показанного в Таблице I, достигая, возможно, всего лишь 10% или ниже расчетного теоретического максимума.Очевидно, что эти три специфические биологические проблемы (a – c) необходимо преодолеть, чтобы повысить фактический урожай зеленой водоросли H 2 .
ВЫВОДЫ
Таким образом, опасения по поводу глобального потепления и загрязнения окружающей среды из-за использования ископаемого топлива в сочетании с прогнозами потенциальной нехватки ископаемого топлива к середине 21-го века делают необходимым развитие альтернативных источников энергии, которые являются чистыми, возобновляемыми и экологически чистый.Недавно разработанный протокол с одним организмом, двухэтапным фотосинтезом и производственным протоколом H 2 с зелеными водорослями представляет интерес, потому что значительные количества газа H 2 были впервые образованы, в основном из солнечного света и воды (Melis et al. , 2000). Кроме того, этот метод не влечет за собой образование каких-либо нежелательных, вредных или загрязняющих побочных продуктов и может даже предложить преимущества продуктов с добавленной стоимостью в результате массового выращивания зеленых водорослей. Однако необходимо преодолеть несколько биологических и инженерных проблем, прежде чем эта многообещающая технология станет реальностью.Прежде всего, клеточный метаболизм и базовая биохимия, которые поддерживают этот процесс, должны быть хорошо изучены, и еще предстоит провести много фундаментальных исследований механизма производства H 2 за счет S-депривации.
В конечном итоге появление водорода вызовет технологические разработки во многих областях, включая производство электроэнергии, сельское хозяйство, автомобильную промышленность и другие, пока еще непредвиденные области применения. Это увеличит занятость, будет стимулировать экономику всех стран на Земле и окажет положительное влияние на окружающую среду, в которой почти полностью уменьшится атмосферное загрязнение и уменьшится так называемый парниковый эффект.
Благодарность
Мы хотим поблагодарить Лора Флорина за помощь с рис. 4.
Сноски
-
№1 Эта работа была частично поддержана Калифорнийской энергетической комиссией, Программой малых грантов энергетических инноваций (грант № 51235A / 99–01–33), Калифорнийской сельскохозяйственной экспериментальной станцией и Deutsche Forschungsgemeinschaft (грант № Ha 2555). / 1–1).
-
↵ * Автор, ответственный за переписку; электронная почта melis {at} nature.berkeley.edu; факс 510–642–4995.
-
www.plantphysiol.org/cgi/doi/10.1104/pp.010498.
- Поступила 5 июня 2001 г.
- Принята 19 августа 2001 г.
Simple English Wikipedia, бесплатная энциклопедия
Dinobryon , колониальная водоросль из группы Chrysophyceae Водоросли ( Laurencia ) вблизи: «ветки» многоклеточные, их толщина составляет всего около 1 мм. Гораздо более мелкие водоросли прикрепляются к структуре, идущей вверх в нижней правой четверти. Цветение фитопланктона в южной части Атлантического океана у побережья АргентиныВодоросли (одна водоросль, но несколько водорослей) - это тип растений, подобных живым существам, которые могут производить пищу из солнечного света путем фотосинтеза.Изучение водорослей называется фикологией или альгологией.
Термин охватывает целый ряд фотосинтезирующих организмов, многие из которых не связаны между собой. Это полифилетическая группа.
Этот термин объединяет множество различных организмов. Их объединяет только то, что они автотрофны: они используют природные источники энергии и простые неорганические материалы для создания своих форм. Как несосудистые растения они не имеют клеточной и тканевой структуры наземных растений. Это удобный, но очень расплывчатый термин.Лишь в последние годы стало ясно, насколько разные виды водорослей.
Водоросли - это большая и разнообразная группа простых, обычно автотрофных организмов. У некоторых есть одна ячейка, а у других - много ячеек. Самые крупные и сложные морские водоросли называются водорослями. Они похожи на растения и «просты», потому что в них отсутствуют многие отдельные органы, обнаруженные у наземных растений. По этой причине их , а не классифицируются как растения.
Хотя прокариотические цианобактерии (ранее называвшиеся сине-зелеными водорослями ) были включены в старые учебники как «водоросли», сейчас их нет. [1] Термин водоросли теперь используется для обозначения эукариотических организмов. [2] Все настоящие водоросли имеют ядро внутри мембраны и хлоропласты внутри одной или нескольких мембран. [1] [3] Однако водоросли определенно являются , а не монофилетической группой, [1] , поскольку не все они происходят от общего предка водорослей. [4] Современные систематики предлагают разделить их на монофилетические группы, но не все согласны с тем, как это сделать. [4] [5]
Водоросли не имеют той же структуры, что и наземные растения, например листья, корни и другие органы. Почти у всех водорослей есть части, которые фотосинтезируют так же, как цианобактерии. Они производят кислород, в отличие от других фотосинтезирующих бактерий, таких как фиолетовых и зеленых бактерий. Некоторые одноклеточные виды используют только внешние источники энергии и не имеют фотосинтетических частей или не имеют их.
Окаменелые нитчатые водоросли из бассейна Виндхья датируются 1 годом.6–1,7 миллиарда лет назад. [6]
Виды водорослей [изменить | изменить источник]
Ниже приведены некоторые важные виды водорослей. Список не полный.
- Зеленые водоросли: их считают растениями, потому что они используют тот же тип хлорофилла, что и зеленые растения. Предполагается эволюционная связь между зелеными водорослями и зелеными растениями.
- Красные водоросли: используют красный пигмент для улавливания энергии солнечного света, поэтому считается, что они эволюционировали отдельно от зеленых растений.
- Бурые водоросли: используют хлорофилл а, но имеют несколько других биохимических отличий. Также не считается зеленым растением.
- Желто-зеленые водоросли: Xanthophyceae.
- Золотые водоросли: Chrysophyceae.
Экология [изменить | изменить источник]
Водоросли обычно встречаются во влажных местах или в воде, а также на суше и в воде. Однако водоросли на суше обычно незаметны и гораздо чаще встречаются во влажных тропических регионах, чем в засушливых. У водорослей нет сосудистых тканей и других приспособлений для жизни на суше, но они могут переносить сухость и другие условия в симбиозе с таким грибком, как лишайник.
Различные виды водорослей играют важную роль в водной экологии. Микроскопические формы, обитающие во взвешенном состоянии в толще воды, называются фитопланктоном. Они обеспечивают пищевую базу для большинства морских пищевых цепочек. Келп растет в основном на мелководье в морской воде. Некоторые из них используются в пищу для людей или собираются для агара или удобрений. Келп может расти в больших насаждениях, называемых лесами ламинарии. Эти леса предотвращают некоторый ущерб от волн. В них обитает много разных видов, в том числе морские ежи, каланы и морские ушки.
Некоторые водоросли могут нанести вред другим видам. Некоторые водоросли могут сильно размножаться и вызывать цветение водорослей. Эти водоросли могут вырабатывать защитные токсины, убивающие рыбу в воде. Динофлагелляты представляют собой соединение, превращающее плоть рыбы в слизь. Затем водоросли поедают эту питательную жидкость.
Симбиоз [изменить | изменить источник]
Nudibranch Pteraeolidia ianthina содержит водоросли, которые фотосинтезируют и обеспечивают моллюсков пищей.Водоросли развили ряд симбиотических партнерских отношений с другими организмами.Самый известный из них - лишайники, похожие на растения, каждый из которых образован грибком с водорослями. Это очень успешная форма жизни, и известно двадцать тысяч «видов». Во всех случаях лишайник совершенно отличается по внешнему виду и образу жизни от того или иного компонента; возможно, это наиболее полный из известных симбиозов. Оба компонента получают выгоду от доступа к нишам с низкой питательной ценностью, в которых встречаются лишайники.
Менее известны взаимоотношения водорослей с животными. Кораллы, строящие рифы, - это в основном социальные книдарийские полипы.Кораллы зависят от света, потому что водоросли - важные партнеры, и им нужен свет. Кораллы имеют развитые структуры, часто похожие на деревья, которые обеспечивают водорослям максимальный доступ к свету. Коралл ослабляет клеточные стенки водорослей и переваривает около 80% пищи, синтезируемой водорослями. Отходы кораллов являются источником питательных веществ для водорослей, поэтому, как и в случае с лишайником, оба партнера получают выгоду от ассоциации. Водоросли представляют собой золотисто-коричневые жгутиковые водоросли, часто принадлежащие к роду Symbiodinium .Любопытной особенностью партнерства является то, что коралл может выбросить водоросли в трудные времена и вернуть их позже. Выброс водорослей-партнера называется обесцвечивание , потому что коралл теряет свой цвет. [7] p200
Другие виды Cnideria, такие как морские анемоны и медузы, также содержат водоросли. Медузы с водорослями ведут себя так, что их партнеры получают лучший свет днем, а ночью спускаются на глубины, где вода богата нитратами и буреет от гниения.Морские слизни и моллюски также хорошо известны тем, что укрывают водоросли. Обе группы - моллюски. Морские слизни пасутся на кораллах, и они того же цвета, что и кораллы, которые они пасутся. Они способны отделять водоросли от перевариваемых тканей полипа. Клетки водоросли перемещаются к его щупальцам, где они продолжают жить. В остальном беззащитный слизняк получает и маскировку, и питание. [7] p204 Гигантский моллюск удерживает водоросли в своей мантии, которая раскрывается, когда моллюск открыт. Цветная мантия имеет места, где кожа прозрачна, и действует как линза, концентрируя свет на водорослях под ней.Когда водорослей становится слишком много, моллюски их переваривают. [7] p203
В различных других группах морских беспозвоночных есть члены, находящиеся в симбиозе с водорослями. Две такие группы - плоские черви (Platyhelminths) и многощетинковые черви (Annelids).
- ↑ 1,0 1,1 1,2 Nabors, Murray W. (2004). Введение в ботанику . Сан-Франциско, Калифорния: Pearson Education, Inc.
- ↑ Allaby, M ed.(1992). «Водоросли». Краткий ботанический словарь . Оксфорд: Издательство Оксфордского университета. CS1 maint: дополнительный текст: список авторов (ссылка)
- ↑ Round, Frank Eric 1981. Экология водорослей . Издательство Кембриджского университета.
- ↑ 4,0 4,1 Патрик Дж. Килинг (2004). «Разнообразие и эволюционная история пластид и их хозяев». Американский журнал ботаники . 91 : 1481–1493. DOI: 10.3732 / ajb.91.10.1481.
- ↑ Парфри, Лаура Вегенер; и другие. (2006). «Оценка поддержки текущей классификации эукариотического разнообразия». PLoS Genet . 2 (12): e220. DOI: 10.1371 / journal.pgen.0020220. PMC 1713255. PMID 17194223.
- ↑ Бенгтсон С., Беливанова В., Расмуссен Б., Уайтхаус М. 2009. Спорные «кембрийские» окаменелости Виндхьяна реальны, но старше их более чем на миллиард лет. PNAS 106 : 7729-34.
- ↑ 7.0 7,1 7,2 Аттенборо, Дэвид. 1995. Частная жизнь растений . BBC, Лондон. Глава 5 Совместная жизнь.
PPT - Introduction to the Classification of Green Algae PowerPoint Presentation
Introduction to the Classification of Green Algae Brian Wysor, Ph.D. Университет Роджера Вильямса
Классификация • Со времен Линнея было описано более 20 классов зеленых водорослей • Многие критерии использовались для определения классов • Морфология • Ультраструктура • + митоз, деление клеток, цитокинез, жгутиковый аппарат • Образцы жизненного цикла • Молекулярный • В первую очередь гены рРНК, rbcL • Неоднозначность остается, но появляется консенсус
Классификация: морфологическая концепция • Предпосылка • Примитивные одноклеточные клетки со временем эволюционировали в структурную сложность • Морфологическое расхождение различает группы Фото Хуанг Су-фан.Pröschold & Leliaert. 2007. Сифонокладозная паренхиматозная эволюция структурной сложности
Проблемы с морфологической концепцией • Селективное давление окружающей среды может способствовать конвергенции • Биологически / генетически различные виды кажутся похожими Charophyceae Chlorophyceae Ulvophyceae 4 Рис. 16.18 Graham et al. 2008.
Морфологическая конвергенция с классом Класс: Ulvophyceae Ulvophyceae Отряд: Ulvales Ulotrichales Род: UlvariaMonostroma
Chlorophyta: морфологическая конвергенция • Как могут быть классифицированы разные образцы зеленых водорослей, если они могут быть классифицированы по образцу конвергенции. морфологии?
Ультраструктурная концепция • Mattox & Stewart (1984) предложили новую таксономию на уровне классов на основе базальной ориентации тела • Первое излучение зеленых водорослей произошло среди одноклеточных жгутиконосцев • Многочисленные древние линии жгутиконосцев диверсифицировались в современные формы на разных этапах эволюции путей, но многие из них имеют схожие морфологические результаты • Выявлено 4 различных паттерна базального тела
Ориентация базального тела Наследственное состояние Ulvophyceae, Trebouxiophyceae, Charophyceae, Drier Green Algae Chlorophyceae Chlorophyceae
Molecular Concept (McCourt & Lewis 2004, Working Classification) молекулярные данные подтверждают признание двух типов и плохо различимые ранние расходящиеся жгутиконосцы, неофициально известные как Prasinophyceans Viridoplantae или Chlorobionta Chlorophyta Streptophyta (Charophyceae + Drier Green Algae)
SSU rDNA Molecular Concept (Pröschold & Leliaert recophyta • с низкой опорой BS (без жирных линий) • Плохо расслоение Prasinophycean, класс 11
Класс Ulvophyceae • Большинство макроскопических морских зеленых водорослей принадлежат к классу Ulvophyceae • Ulvophyceae определяется на основе набора плейсиоморфных признаков • Закрытый митоз, стойкий шпиндель, furro крыло, CCW, жгутиковые клетки с чешуей • Отсутствие четко определенных общих производных признаков привело к неопределенным отношениям внутри класса
Класс Ulvophyceae McCourt & Lewis.2004. • Обычно различают 5 (или 6) порядков • жгутиковый аппарат • зооспорангиальные и гаметангиальные структуры • жизненный цикл • Порядки обычно подтверждаются молекулярным анализом, но отношения между порядками в значительной степени неопределенны.
Molecular Concept (Lopez- Bautista et al. 2002)
Субординальная классификация • Классификация на уровне родов и видов часто основана на морфологии, которая может быть проблематичной • Фенотипическая пластичность • Морфологическая конвергенция • Для решения этих проблем требуются комплексные подходы • Секвенирование ДНК (и дистанционная кластеризация) - это начало, которое может привести к распознаванию новых морфологических, физиологических или биологических особенностей
Z-клады и биологические виды, выведенные из структуры ITS2 2 ° (см. Coleman 2009) увеличение потенциальных партнеров, снижение сексуальной совместимости и продуктивности зигот • Организм ms, которые отличаются даже одним CBC в спирали III, совершенно неспособны к скрещиванию • Идентичность для всего ITS2 коррелирует со значительным потенциалом межпородного скрещивания.
Цитированная литература • Coleman, A. 2009. Существует ли молекулярный ключ к уровню «биологических видов» у эукариот? Гид по ДНК. Мол. Филоген. Evol. 50: 197-203. • Graham, L.E., Graham, J.M., & Wilcox, L.W. 2008. Водоросли, 2-е изд., Бенджамин Каммингс, Сан-Франциско, 616 с. • Mattox, K.R. И Стюарт, К. 1984. Классификация зеленых водорослей: концепция, основанная на сравнительной цитологии. Стр. 29–72, в Ирвине, D.E.G. И Джон Д. (ред.), Систематика зеленых водорослей.Ассоциация систематиков, Special Vol. 27, Academic Press, Лондон. • Маккорт Р. и Льюис Л. 2004. Зеленые водоросли и происхождение наземных растений. Am. J. Bot. 91: 1535-1556. • Pröschold, T. & Leliaert, F. 2007. Систематика зеленых водорослей: конфликт классического и современного подходов. Стр. 123-153 В Броди, Дж. И Льюисе, Дж. Распутывание водорослей: прошлое, настоящее и будущее водорослевой систематики. CRC press, Boca Raton, 376 p. • Ван ден Хук, К., Манн, Д.Г., и Янс, Х.М. (1995) Водоросли.Введение в психологию. Издательство Кембриджского университета, Кембридж.
Эта презентация является вкладом Панамериканского института перспективных исследований
% PDF-1.4 % 49 0 объект > endobj 51 0 объект > поток 1998-01-09T10: 40: 44Z2020-12-27T19: 19: 47-08: 002020-12-27T19: 19: 47-08: 00 Acrobat Distiller Command 3.01 для Solaris 2.3 и более поздних версий (SPARC) uuid: 736925be-1dd2- 11b2-0a00-8909275dc400uuid: 736925c1-1dd2-11b2-0a00-810000000000application / pdf конечный поток endobj 11 0 объект > endobj 1 0 obj > / Font> / ProcSet [/ PDF / Text] >> / Type / Page >> endobj 13 0 объект > / Font> / ProcSet [/ PDF / Text / ImageB] / XObject >>> / Type / Page >> endobj 17 0 объект > / Font> / ProcSet [/ PDF / Text / ImageB] / XObject >>> / Type / Page >> endobj 21 0 объект > / Font> / ProcSet [/ PDF / Text / ImageB] / XObject >>> / Type / Page >> endobj 24 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Type / Page >> endobj 28 0 объект > / Font> / ProcSet [/ PDF / Text / ImageB] / XObject >>> / Type / Page >> endobj 31 0 объект > / Font> / ProcSet [/ PDF / Text] >> / Type / Page >> endobj 77 0 объект [80 0 R] endobj 78 0 объект > поток HWr8} WQ ޒ8% ۉ qYJR [D! 8 @ ҃Nv7N \, = g- ex {hꗌ | al4 ч | W ^^ L4eMha42 ϧ "F n ^ X҈1Kʟ ~ (fœtXkWnu, 7eYsIUӯ * | -WsUmey7Q1X ~ S4OasCnZqJ = 庬 YIGaM ݫ} IjGh_ + V;` wCq & lVVϪ5wnQm ~ 1 00 "@