 Категории
Категории Корзина
Корзина Информация
Информация Хиты продаж
Хиты продаж






Особенности строения органов чувств у рыб
Органы чувств рыб. Строение органов зрения, слуха, боковой линии
К органам чувств рыбы относятся: зрение, слух, боковая линия, электрорецепция, обоняние, вкус и осязание. Разберем каждое по отдельности.
Орган зрения
Зрение – один из основных органов чувств у рыб. Глаз состоит из округлой формы хрусталика, имеющего твердую структуру. Находится вблизи роговицы и позволяет видеть на расстояние до 5м в состоянии покоя, максимальное зрение достигает 10-14м.
 Строение глаза рыбы
Строение глаза рыбыХрусталик улавливает множество световых лучей, позволяя видеть в нескольких направлениях. Часто глаз имеет возвышенное положение, таким образом, в него попадают прямые лучи света, косые, а также сверху, снизу, с боков. Это значительно расширяет поле зрения рыб: в вертикальной плоскости до 150°, а в горизонтальной – до 170°.
Зрение монокулярное – правый и левый глаз получает отдельное изображение. Глаз состоит из трех оболочек: склера (ограждает от механических повреждений), сосудистой (поставляет питательные вещества), и ретинальной (обеспечивает световосприятие и цветоощущение за счет системы палочек и колбочек).
Орган слуха
Слуховой аппарат (внутреннее ухо или лабиринт) расположен в задней части черепной коробки, включает два отделения: верхний овальный и круглый нижний мешочки. В овальном мешочке расположены три полукружных канала – это орган равновесия, внутри лабиринта течет эндолимфа, с помощью выводного протока соединяется у хрящевых рыб с окружающей средой, у костных — заканчивается слепо.
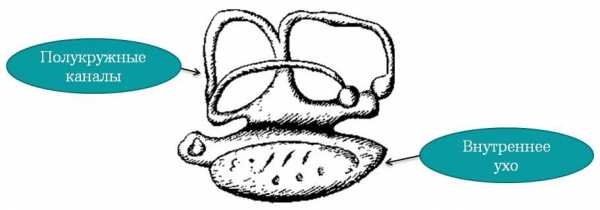 Орган слуха у рыб совмещен с органом равновесия
Орган слуха у рыб совмещен с органом равновесияВнутреннее ухо делится на три камеры, в каждой находится отолит (часть вестибулярного аппарата, который реагирует на механическое раздражение). Внутри уха заканчивается слуховой нерв, образуя волосковые клетки (рецепторы), при изменении положения тела раздражаются эндолимфой полукружных каналов и помогают сохранять равновесие.
Восприятие звуков осуществляется за счет нижней части лабиринта – круглого мешочка. Рыбы способны улавливать звуки в диапазоне 5Гц – 15кГц. К слуховому аппарату относятся боковая линия (позволяет услышать низкочастотные звуки) и плавательный пузырь (выступает как резонатор, соединён с внутренним ухом посредством Веберового аппарата, состоящего из 4 косточек).
Рыбы близорукие животные, передвигаются часто в мутной воде, с плохим освещением, некоторые особи обитают в морских глубинах, куда свет не достает вовсе. Какие же органы чувств и как позволяют ориентироваться в воде при таких условиях?
Боковая линия
Прежде всего – это боковая линия – основной орган чувств у рыб. Представляет собой канал, который идет под кожей вдоль всего тела, в области головы разветвляется, образуя сложную сеть. Имеет отверстия, через которые связывается с окружающей средой. Внутри расположены чувствительные почки (рецепторные клетки), которые воспринимают малейшие изменения вокруг.
Так они могут определять направление течения, ориентироваться на местности ночью, ощущать движение других рыб, как в стае, так и приближающихся к ним хищников. Боковая линия оснащена механорецепторами, они помогают водным жителям уворачиваться от подводных камней, инородных предметов, даже при плохой видимости.
Боковая линия может быть полной (располагается от головы до хвостовой части), неполной, а может быть вовсе заменена на другие развитые нервные окончания. При травмировании боковой линии рыба уже не сможет долго существовать, что свидетельствует о важности данного органа.
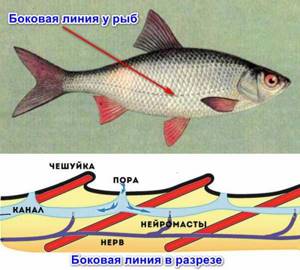 Боковая линия рыб — главный орган ориентации
Боковая линия рыб — главный орган ориентацииЭлектрорецепция
Электрорецепция – орган чувств хрящевых рыб и некоторых костистых (электрический сом). Акулы и скаты ощущают электрические поля с помощью ампул Лоренцини – небольшие капсулы заполненные слизистым содержимым и выстланы специфическими чувствительными клетками, находятся в области головы и сообщаются с поверхностью кожи при помощи тонкой трубки.
Очень восприимчивы и способны ощущать слабые электрические поля (реакция возникает при напряжении в 0,001 мКв/м).
Так электрочувствительные рыбы могут выследить жертву, скрытую в песке, благодаря электрическим полям, которые создаются при сокращении мышечных волокон во время дыхания.
Боковая линия и электрочувствительность – это органы чувств характерны только для рыб!
Орган обоняния
Обоняние осуществляется при помощи ресничек, расположенных на поверхности специальных мешочков. Когда рыба чует запах, мешочки начинают двигаться: сужаться и расширятся, улавливая пахучие вещества. Нос включает 4 ноздри, высланные множеством чувствительных клеток.
Своим нюхом легко находят пищу, сородичей, партнера на период нереста. Некоторые особи способны подавать сигналы об опасности выделяя вещества, к которым чувствительны другие рыбы. Считают, что обоняние для водных жителей важнее зрения.
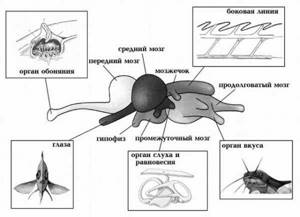 Органы чувств у рыб
Органы чувств у рыбОрганы вкуса
Вкусовые рецепторы рыб сосредоточены в ротовой полости (ротовые почки), и ротоглотке. У отдельных видов (сом, налим) встречаются в области губ и усов, у сазанов — по всему телу.
Рыбы способны распознавать, как и человек, все вкусовые характеристики: соленое, сладкое, кислое, горькое. С помощью чувствительных рецепторов рыба может отыскать необходимую пищу.
Осязание
Рецепторы осязания расположены у хрящевых рыб на участках тела не покрытых чешуей (брюшная область у скатов). У костистых чувствительные клетки разбросаны по всему телу, основная масса сосредоточена на плавниках, губах — дают возможность ощущать прикосновения.
Особенности органов чувств у костистых и хрящевых
Косные рыбы имеют плавательный пузырь, который воспринимает более широкий диапазон звуков, у хрящевых он отсутствует, также у них идет не полное разделение внутреннего уха на овальный и круглый мешочки.
Цветное зрение свойственно костистым, поскольку в их сетчатке находятся и палочки, и колбочки. Зрительный орган чувств хрящевых включает лишь палочки, которые не способны различать цвета.
У акул очень острый нюх, намного больше развита передняя часть мозга (обеспечивает обоняние), чем у других представителей.
Электрические органы – особые органы хрящевых рыб (скатов). Используются для защиты, нападения на жертву, при этом генерируются разряды мощностью до 600В. Могут выступать в качестве органа чувств – образуя электрическое поле, скаты улавливают изменения при попадании в него посторонних тел.
Органы чувств у рыб: описание, особенности, факты
Человек привык экстраполировать собственные ощущения и на окружающий мир. Мы подсознательно полагаем, что представители животного мира, в том числе, и ихтиофауны, воспринимают окружающую среду по аналогии с человеком. А между тем рыбы хоть располагают органами чувств, подобными нашим, реагируют на внешние раздражители несколько иначе. Более того, в их сенсорном арсенале есть и не присущие человеку способы восприятия.
Давайте разбираться, как же чувствуют и воспринимают внешние раздражители представители ихтиофауны. Это поможет нам понять, как правильно вести себя на рыбалке, что позволительно делать, а от чего лучше воздержаться. Ориентируясь на эту информацию, можно правильно выбрать как тактику ловли, так и наиболее результативную в конкретных условиях приманку. Да и вообще, предполагаемого оппонента нужно знать досконально!
Что чувствуют рыбы?
Рыбы – удивительные существа древнейшего происхождения. Наверняка все вы знаете, что жизнь зародилась в Мировом океане. Со временем часть обитателей в ходе эволюции покинула водную «колыбель», но предки современных рыб предпочли остаться в привычной среде. Шли тысячелетия, наземные жители все более совершенствовались и образовывали новые классы, семейства и виды. Затронула эволюция и рыб, усовершенствовав их механизмы приспосабливаемости к водной стихии и сенсорику.
Эта приманка обеспечивает богатый улов даже при плохом клеве!
ПодробнееПоговорим о лучеперых рыбах – именно они составляют основу современной ихтиофауны (подавляющее большинство ныне живущих обитателей гидросферы относится именно к этому обширнейшему классу), а конкретнее – об их сенсорике.
Известно, что человек обладает пятью органами чувств. Это зрение, слух, обоняние, осязание, вкус. Сходные органы восприятия есть и у большинства рыб, но устроены они несколько иначе. Более того: представители ихтиофауны обладают и абсолютно уникальными рецепторами, аналогов которым у наземных животных нет вообще. Это боковая линия, которая по праву может считаться главным рецептором большинства рыб. Некоторые представители ихтиофауны обладают и седьмым чувством, позволяющим улавливать возмущения электрополей. Видите, сколько интересного? А теперь обо всем по порядку.
Зрение
В повседневной жизни мы более всего привыкли полагаться на зрение: львиную долю информации мы получаем при помощи глаз. У рыб сложилась несколько иная ситуация: это чувство играет в их жизни не столь важную роль, хотя и абсолютно слепых среди них маловато (за исключением самых глубоководных видов).
Глаза рыб устроены привычным для нас образом: радужка, зрачок (в отличие от нашего – у большинства видов не реагирующий на степень освещенности), хрусталик, сетчатка. Сетчатка состоит из фоторецепторов (палочек и колбочек), которые и отвечают за качество изображения.
Давайте вкратце пробежимся по основным характеристикам зрения среднестатистического обитателя водоема:
- Угол зрения. Глаза у большинства пресноводных рыб расположены по бокам головы, подвижны, но зависимы друг от друга (независимо вращать зрительными рецепторами подобно хамелеону рыбы не умеют). Зона бинокулярного зрения у них невелика (порядка 30о), зато угол монокулярного обзора у них поистине завидный – до 170о, что позволяет им видеть хищника или пищу практически всегда. Основная слепая зона располагается за хвостом рыбы, поэтому наиболее уязвима она «с тыла». Если учитывать коэффициент преломления света на границе водной и воздушной среды, а также особенности органов зрения рыб, можно смело утверждать, что объекты, расположенные на берегу под углом менее 45о, для них неразличимы. То есть, сидящего на берегу или в лодке человека рыба, скорее всего не увидит, а вот стоящего – запросто.
- Острота. На дальних дистанциях рыба, скорее, чувствует объекты, нежели видит. Крупные объекты рыбы различают с расстояния 10-12 метров, но четко дифференцируют их с более близкого расстояния. То есть, внешний вид приманки имеет значение лишь на расстоянии 2-5 метров, а в мутной воде или в темень – и того меньше.
- Цветоразличение. Большинство рыб неплохо различает цвета, во всяком случае, чистой палитры (в сетчатке присутствуют колбочки, отвечающие за цветоразличение). Однако глубокосумеречные и ночные виды (те же судак и налим) практически полные дальтоники: в их сетчатке почти нет колбочек. Зато они лучше различают предметы в темноте, особенно контрастные по оттенку. То есть, при ловле того же судака цветовые нюансы приманки можно сбрасывать со счетов, но светлые тона, выделяющиеся из окружающейся среды, в приоритете.
Слух
Даже человек, далекий от рыбалки, знает, что шуметь на берегу крайне не рекомендуется. Однако водные жители прекрасно чувствуют себя под мостами, где организовано движение большегрузов и в прудах вблизи населенных пунктов. Быть может, они ничего не слышат? Нет, просто к фоновым звукам представители ихтиофауны привыкают и никак не реагируют на обыденные шумы.
Органы слуха у рыб устроены принципиально иначе, чем у нас, и чувствуют звуковые колебания они несколько по-другому. Ушей в привычном для нас понимании у обитателей водоемов нет, но имеется лабиринт (внутреннее ухо), представленный тремя полукружными каналами, заполненными жидкостью. В них имеются белые камешки (отолиты), которые вибрируют при воздействии акустических волн.
Плавательный пузырь является дополнительным резонатором, а боковая линия позволяет воспринимать недоступные для нашего слуха низкочастотные колебания. Но и верхний порог восприятия у рыб ниже: хищники не слышат звуки высотой свыше 500 Гц, белая рыба – 2 кГц. При этом мы способны воспринимать волны частотой до 20 кГц!
Водная среда прекрасно проводит звук – в 4,5 раза лучше, чем воздух. Даже негромкий звук разносится за многие километры, чем и пользуются любители ловить сомов «на квок» (специальным инструментом ударяют по воде, порождая звуки, привлекательные для речных гигантов) и приверженцы искусственных приманок с «погремушками». Однако на границе воздуха и воды трафик весьма велик, поэтому достаточно громкий (но не резкий) звук в воде практически не слышим. Это значит, что на берегу можно вести себя более-менее свободно (в пределах разумного), но в воде и на льду нужно соблюдать осторожность: рыбы ощущают непривычные для них акустические колебания и спешат уйти в безопасное место.
Обоняние
У большинства пресноводных рыб имеются внешние и внутренние органы обоняния, на которые они полагаются в большей степени, чем на глаза. Они пропускают воду через ноздри (обычно их две) и при помощи соответствующих рецепторов, расположенных в обонятельных складках, анализируют растворенные в ней микродозы различных веществ. Вы не поверите, чувство обоняния у некоторых видов рыб развито не хуже, чем у признанных «нюхачей» сухопутного мира, собак.
Несомненным чемпионом в этой номинации является речной сом, но и прочие известные нам представители ихтиофауны различают запахи куда лучше, чем мы с вами. Благодаря этому чувству они находят пищу, выбирают партнеров в период размножения, спасаются от природных врагов.
Акула чувствует запах крови, а карп – аромат свежей макухи за несколько километров. Это используется рыболовами при подборе приманок: широко применяются привлекающие рыбу ароматические добавки, именуемые аттрактантами или активаторами клева. Но никакой аттрактант не способен забить непривлекательный для рыбы запах, поэтому прокисшие, залежавшиеся или испорченные иным образом приманки под запретом.
Высокая чувствительность к запахам – это еще что! Рыбы способны анализировать даже химический состав воды, степень ее насыщенности кислородом и прочие параметры! Это позволяет тем же лососям безошибочно находить реки, в которых они появились на свет, а угрям – прицельно устремляться в Саргассово море за тысячи километров. Правда, это чувство нельзя назвать обонянием в привычном нам смысле этого слова, но какая разница: рыбы успешно используют его в процессе выживания.
Вкус
Рыба может получить практически исчерпывающуюся информацию о потенциально съедобном объекте, руководствуясь исключительно обонянием. Однако и вкус играет немаловажную роль в пищевом поведении большинства представителей ихтиофауны. Рыбы отлично различают вкусы: горький, сладкий, кислый, соленый. Более того: они ощущают вкусовые оттенки гораздо более тонко, чем мы с вами.
При приближении к потенциально съедобному объекту рыбы обычно исследуют его при помощи внешних вкусовых рецепторов, расположенных на губах, усиках, жабрах, голове, а иногда даже плавниках. Те же карпы долго «смакуют» предложенную пищу, прогоняя через жабры воду с растворенными в ней питательными веществами.
Разумеется, если рыба очень голодна, она пренебрегает исследованием лакомства при помощи внешних органов чувств, сразу хватая пищу и лишь потом анализируя вкус при помощи рецепторов, расположенных уже во рту (как у нас). Если вкус приманки не коррелирует с издаваемым ей запахом, вполне возможно, что представительница ихтиофауны выплюнет предложенное лакомство с негодованием.
Вкусовые предпочтения различных рыб определяются составом их основной кормовой базы в конкретный сезон. Среди рыболовов бытует поверье, что карпы любят сладенькое. Однако исследования показали, что карпы более неравнодушны к кислому, что объясняет успех приманок с добавлением фруктовых и ягодных компонентов. А вот амур, плотва и елец действительно предпочитают пищу с большим содержанием сахарозы. Голавль же любит небольшое добавление пикантной горечи: она присутствует в хитине воздушных насекомых, коими он преимущественно питается. Отсюда вывод: при подборе приманки нужно ориентироваться на особенности кормовой базы конкретного водоема, не забывая и о сезонности.
Осязание
Рыбы, как и мы с вами, способны чувствовать прикосновения, боль, температурные колебания. Это обязательно для обеспечения выживания: рыба вынуждена реагировать на боль и колебания температуры. Помните, как уколовшись о крючок, она выпускает сомнительную добычу изо рта. Правда, нервная система у представителей ихтиофауны не отличается высокой степенью развития, посему болевой порог у них низок.
Соответствующие тактильные и температурные рецепторы расположены по всему телу рыбы: от верхних слоев кожи и плавников до слизистых оболочек и мягких тканей. Особую роль в процессе выживания играют терморецепторы, чутко реагирующие на изменения температуры. Дело в том, что рыбы – хладнокровные существа, и они не умеют поддерживать температуру тела подобно нам с вами. Следовательно, они должны каким-то образом приспосабливаться.
Для каждого вида рыб характерен свой комфортный температурный диапазон. Например, при температуре ниже 8оС карась и карп уже впадают в состояние, близкое к анабиозу, в то время, как форель или налим чувствуют себя комфортно и в куда более холодной воде (только бы не замерзала). Зато хладолюбивые рыбы испытывают настоящий термошок при температуре воды свыше 20оС, когда вышеупомянутые карповые резвятся вовсю. И это нужно учитывать при выборе времени, стратегии и тактики ловли рыб различных видов.
Боковая линия
Боковая линия – поистине уникальный орган чувств, присущий исключительно представителям ихтиофауны и некоторым земноводным, практически постоянно обитающим в воде. Видели у рыб этакую линию, проходящую по бокам с обеих сторон приблизительно посередине тела? Этот главный канал буквально усеян рецепторами, отвечающими за сейсмосенсорику. Внешние выводы их располагаются на чешуйках и голове, а нейроны связывают их с головным мозгом.
Боковая линия может быть сплошной и хорошо различимой, может прерываться, а у некоторых рыб и вовсе располагаться в нетипичном месте. Например, у сельди органы сейсмосенсорики расположены на голове.
В ходе экспериментов замечено, что даже лишившись зрения, многие рыбы вполне неплохо себя чувствуют. Боковая линия позволяет им различать малейшие колебания в среде, в том числе – и тонко улавливать изменение направления тока воды. Вследствие этого рыбы не натыкаются на препятствия даже в полной темноте или мутной воде, распознают характерные колебания, издаваемые потенциальной добычей (например, копошение мотыля в придонном иле), чутко реагируют на приближение хищника.
Например, если лишить зрения щуку, она вполне успешно продолжит охотиться, полагаясь на иные органы чувств. А вот если разрушить ей и боковую линию, хищница, даже очень проголодавшись, не отреагирует на резвящуюся рядом лакомую рыбешку.
Вывод: нужно постараться не создавать в воде настораживающих колебаний, то есть не топать на берегу, тихо ходить по льду, соблюдать осторожность в лодке.
Электрорецепция
Некоторые рыбы способны генерировать слабое электромагнитное поле. Подобные органы имеются у электрических угрей, скатов, сомов. Этих рыб окружает электромагнитное поле, и они чутко реагируют на его возмущение, вызванное попаданием различных объектов.
Например, в Ниле проживает поразительная рыба, именуемая длиннорылом или водяным слоном. Но самое удивительное в ней не наличие длинного «носа» подобного слоновьему хоботу, а умение «видеть хвостом» (во всяком случае, так полагают местные жители). В ходе исследований было установлено, что в районе «кормы» у этой рыбы располагается своеобразный природный генератор, а в особом плавнике – орган чувств, отвечающий за электрорецепцию.
Органы электрорецепции водяного слона очень чувствительны: он способен уловить упавшую в воду песчинку. Это позволяет ему успешно избегать рыболовных сетей и прочих объектов, грозящих опасностью.
Интересные факты о рыбах
И теперь, в качестве десерта к публикации, несколько занимательных фактов об органах чувств интересных представителей ихтиофауны:
- Камбала. Эта удивительная рыба на стадии малька мало чем отличается от прочих представителей ихтиофауны. Затем природа начинает придавать ей характерный облик: тело уплощается, а глаза съезжают на одну сторону. Большую часть жизни камбала проводит лежа на дне. Ее чешуя наделена способностью к мимикрии, но утратив зрение, камбала уже не может слиться с окружающей средой. Однако это не мешает ей активно питаться и вполне неплохо себя чувствовать.
- Зеленушка. Основу кормовой базы этой черноморской рыбки составляют моллюски. Если зеленушка потеряет зрение, корма она не найдет и, разумеется, погибнет, так как прочие органы чувств у нее развиты очень слабо.
- Налим. Как-то популяцию налимов, облюбовавшую устье реки Оулуйоки, постиг странный недуг, вызванный специфическими паразитами. Вследствие этого практически треть популяции пресноводной трески ослепла. Однако выглядят слепые налимы неплохо – ничуть не менее упитанными, нежели зрячие собратья. Дело в том, что пресноводная треска ведет преимущественно ночной образ жизни, поэтому привыкла полагаться на иные органы чувств.
- Угорь. Эта удивительная рыба визуально похожа на змею. Обитает она в некоторых водоемах Европейской части России и очень ценится за поистине деликатесное мясо. Удивителен не столько внешний вид угря, сколько его образ жизни. Дело в том, что нерестятся угри исключительно в Саргассовом море, порой преодолевая десятки тысяч километров, включая переползаемые ими участки суши. Каким чувством руководствуются эти рыбы в пути – до сих пор остается загадкой.
- Белуга. Эта занесенная в Красную книгу рыба из семейства осетровых считается крупнейшим представителем пресноводной ихтиофауны (научно доказано, что некоторые особи способны нагулять порядка полутора тонн живой массы). Так вот, ученые полагают, что белуги имеют природные локаторы, наподобие дельфинов и летучих мышей. Да уж, таким громадинам встроенные радары точно не помешают.
Мир рыб прекрасен в своем многообразии. Давайте постараемся, чтобы им восхищались наши внуки и правнуки!
Рыболовы удивляются, почему у меня клюет, а у них нет? Только для вас раскрываю секрет: все дело в чудо-приманке!
ПодробнееВнешнее строение рыб. Органы чувств — урок. Биология, Животные (7 класс).
Сжатое с боков тело рыб имеет обтекаемую форму, что уменьшает сопротивление воды при их передвижении. В нём можно различить голову, туловище и хвост.
Рыбы имеют конечности — непарные и парные плавники.
Непарные — это спинные, хвостовой и анальный (подхвостовой).
Снизу по бокам расположены парные плавники (конечности): передние — грудные и задние — брюшные.
Кожа рыб двухслойная, тонкая, слизистая, покрытая костной чешуёй. Чешуя постоянно растёт, и на ней образуются годовые кольца, по ним можно определить возраст рыбы.
Слизь, выделяемая кожными железами рыбы, обеспечивает ей скольжение, уменьшает трение при плавании.
По бокам тела рыбы находятся органы боковой линии — каналы, лежащие в коже под чешуёй, на дне которых расположены чувствительные клетки, воспринимающие колебания воды. При помощи органа боковой линии рыба воспринимает направление течения и давление воды, наличие препятствий, звуковые колебания.
На голове спереди расположен рот, которым рыба захватывает пищу и втягивает воду, необходимую для дыхания.
Выше рта расположены ноздри, открывающиеся в органы обоняния, с помощью которых рыба воспринимает запахи веществ, растворённых в воде.
Глаза рыб довольно большие, они имеют плоскую роговицу, шарообразный хрусталик. Век нет. Рыбы видят на близком расстоянии и различают цвета.
Органы слуха (внутреннее ухо) расположены по бокам головы внутри черепа.
Рядом с внутренним ухом находится орган равновесия, благодаря которому рыба ощущает положение своего тела, перемещения вверх и вниз.
Источники:
Биология. Животные. 7 кл.: учеб. для общеобразоват. учреждений / В. В. Латюшин, В. А. Шапкин. — М.: Дрофа.Трайтак Д. И., Суматохин С. В. Биология. Животные. 7 класс. — М.: Мнемозина.
Никишов А. И., Шарова И. Х. Биология. Животные. 7 класс. — М.: Владос.
Константинов В. М., Бабенко В. Г., Кучменко B. C. / Под ред. Константинова В. М. Биология. 7 класс. — Издательский центр ВЕНТАНА-ГРАФ.
Иллюстрации:
http://school-collection.edu.ru
Органы чувств рыб: общая характеристика
Все внешние стимулы, доступные для анализа органами чувств рыб, можно разделить на несколько видов согласно их природе. Рыбы могут различать химические стимулы, такие как вкус и запах, механические – например, звук, осязание, колебания внешней среды, а также реагировать на электрические и магнитные поля и, конечно, свет.За восприятие всего этого отвечают специализированные органы чувств. Мы привыкли, что у человека за восприятие сигнала определенной природы ответственен какой-то определенный орган – например, за восприятие света отвечает глаз, а звуковых сигналов – ухо. У рыб же часто бывает затруднительным определить конкретный орган, отвечающий за восприятие конкретных сигналов – сигналы сходной природы могут восприниматься несколькими различными органами. Так, за восприятие сигналов химической природы ответственны три самостоятельные хемосенсорные системы – обоняние, вкус и общее химическое чувство, а за восприятие колебаний водной среды механосенсорные системы – вестибулярная, осязание, слуховая, боковой линии.
Ниже я расскажу о том, какие же сигналы и какими органами воспринимают рыбы, а так же о разнообразии внешнего строения этих органов. Это будет только самая общая информация – подробно о каждой из сенсорных систем я буду говорить в отдельных постах.
Органы чувств рыб
Органы боковой линии. Органы боковой линии, свойственные вообще первичноводным позвоночным (круглоротым, рыбам, многим земноводным), достигают у рыб наибольшего развития.
Обычно они расположены по одной или нескольким линиям, тянущимся вдоль боков туловища и хвостового отдела. Особенного развития они достигают на голове, где образуют сложную сеть разветвленных каналов. У химер и примитивных акул органы боковой линии, имеющие строение чувствительных луковиц, располагаются на дне открытого желобка, у прочих рыб они лежат в замкнутом канале, который сообщается с наружной средой отверстиями, прободающими отдельные чешуи.
Органы боковой линии воспринимают звуки низкой частоты от 5 до 25 герц.
Органы вкуса у рыб располагаются не только в ротовой полости, но и на наружной поверхности тела. Они имеют строение отдельных чувствующих почек на наружной поверхности эпидермиса. Особенно сильного развития достигают вкусовые почки у донных рыб на нижней поверхности головы и туловища и служат для распознавания пищи.
Органы обоняния играют у рыб большую роль при питании. У всех рыб, за исключением двоякодышащих, органы обоняния имеют форму парных мешков со складчатыми стенками и открываются наружу одной или двумя ноздрями. У акуловых рыб они расположены на брюшной стороне головы, у всех костных они передвинуты на бока головы, впереди глаза. У двоякодышащих и кистеперых рыб образуются внутренние ноздри (хоаны), открывающиеся в ротовую полость, как у наземных позвоночных.
Орган слуха представлен только внутренним ухом, и звуковые волны передаются ему непосредственно через ткани. Звуковые колебания от 16 до 13 000 гц воспринимаются нижней частью перепончатого „лабиринта (sacculus и lagena). Скорость звука в воде много больше, чем в воздухе (около 1500 м/сек), и восприятие звуковых колебаний позволяет рыбе хорошо ориентироваться в пространстве, отыскивать пищу и избегать опасности. Хорошо воспринимают звуковые колебания рыбы, имеющие веберов аппарат — цепочку из трех подвижно сочлененных косточек, соединяющих лабиринт с плавательным пузырем (сомовые, карпообразные), у лабиринтовых острота слуха увеличивается благодаря тому, что их воздушная камера граничит с sacculus.
Рыбы не только воспринимают звуковые колебания, но и издают звуки; звуковая сигнализация в жизни рыб имеет большое значение. Многие рыбы издают звуки с помощью мускулов, расположенных на плавательном пузыре, который при этом служит в качестве резонатора. Морские петухи и горбылевые издают характерные звуки, напоминающие хрюканье и барабанный бой. Но в то же время известны рыбы, издающие звуки, хотя плавательный пузырь у них отсутствует (например, бычок-кругляк).
В разное время года и суток рыбы издают звуки с разной степенью интенсивности. Некоторые рыбы издают громкие звуки главным образом в период размножения. Это известно для многих видов, охраняющих икру (морской мичман, бычок-кругляк).
Верхняя часть лабиринта представлена тремя полукружными каналами, которые, соединяясь, образуют расширение (utriculus). В utriculus и sacculus имеются отолиты. У костистых рыб они прикрепляются к волоскам чувствительного эпителия, и при изменении положения изменяется тяга волосков; возникшие таким образом сигналы вызывают рефлекторное движение мышц. Изменение внешнего давления через плавательный пузырь и систему косточек веберова аппарата (у сомовых и карповых рыб) передается слуховому лабиринту и продолговатому мозгу, которые регулируют содержание газов в плавательном пузыре.
Еще интересные статьи по теме:
Основные органы чувств у рыб
Приобретайте качественные товары по доступным ценам в лучших рыболовных интернет магазинах. Делайте подарки себе и своим близким!
Мы в социальных сетях - подписывайтесь на нас в Facebook, Youtube, Вконтакте и Instagram. Будьте в курсе последних новостей сайта.
Оглавление:Рыболовы, которые много времени уделяют своему увлечению, знают от чего зависит поведение тех или иных рыб. Они изучают повадки мирных и хищных рыб. На основе имеющихся знаний выстраивают тактику ловли различных рыб в разное время года.
В данной статье мы подробно рассмотрим, какие органы чувств у рыб являются главными. Как они влияют на ее поведение и что должен знать рыболов, чтобы более уверенно чувствовать себя на водоеме.
Какие органы чувств есть у рыб?
Органами чувств рыб, которые прямым образом влияют на их поведение в различных ситуациях, являются органы зрения, осязание, обоняние, боковая линия органы слуха. При помощи них рыбы определяют температуру окружающей среды, различают цвета, определяют качество воды и вкусы, апахи различных предметов и объектов.
Большое значение на поведения рыб оказывают тактильные ощущения. Тело рыбы покрыто множеством нервных окончаний, благодаря которым рыбам легко ориентироваться в водном мире.
Вкусовые ощущения и обоняние у рыб
За вкусовые ощущения и обоняние у рыб отвечают два отверстия на лобной части головы - ноздри. Как у человека нос, так у рыб эти дырочки служат для определения запахов и вкусов разных предметов. Обонятельный орган у рыб позволяет им безошибочно определять путь к нерестилищу или находить растительную и животную пищу в воде.
Лучше всего обоняние развито у тех рыб, которые любят вести активный образ жизни в ночное время и в местах со слабой освещенностью. К таким рыбам относят налима, сома, леща, угря и в какой-то степени сазана. Они очень хорошо различают соленые, сладкие, кислые и горькие запахи.
Вкусовые рецепторы находятся внутри рта, в районе челюстей и на усах рыб. Если посмотреть, как развит ротовой орган у лещей, сазанов, как они легко находят пищу, то многое становится понятным. К примеру, некоторые виды рыб откладывают игру вдали от основных мест обитания. К таковым относятся, в первую очередь, угри, лососи, плотва, вобла и караси в какой-то мере.
Что интересно, мальки, вылупляясь из икринок не могут знать, где их естественная среда обитания. Но они быстро находят дорогу за многие сотни километров и оказываются в кругу своих родственников. Ярким примером являются лососи, которые рождаются в море, а потом с большой скоростью направляются домой. Причем находят именно ту реку, где живут лососи. Определяют свою родную стихию по составу воду, по ее вкусу. За это и отвечают органы обоняния. С их помощью рыбы безошибочно определяют для себя ту воду, в которой они с наибольшим комфортом могли бы жить.
То же самое можно сказать и об угрях. Эти рыбы плывут за тысячу километров, чтобы отложить икру. И они без труда находят родную речку. Не удивительно, что ночью эти рыбы легко находят червей и другую пищу. Зрение по большому счету им необходимо по стольку-поскольку.
Функция органов вкуса и обоняния заключается в определении кислотности среды и количества кислорода в воде. Именно поэтому тот же лещ или густера не будут жить в илистых участках водоема. Такой состав воды им не подходит в отличие от карася или карпа. Грубо говоря, если искусственно поместить рыб в неестественную для них среду, то они там не приживутся. Ноздри у рыб не соединены с носоглоткой. Поэтому вкусовые рецепторы разбросаны по всему телу: на усиках, плавниках и жабрах, а также на коже.
Красноречивыми являются примеры сома и налима, которые очень любят охотиться в ночное время. У этих рыб очень хорошо развиты органы, отвечающие за вкусы и запахи: усы, плавники и ноздри. Налим при помощи усов и плавников без труда находит пищу в холодное и темное время. Сом делает точно также, только в теплое время года.
Большинство мирных и хищных рыб улавливают электропроводимость воды. За эту способность отвечают ямки, расположенные на теле.
Рыбакам следует четко понимать, что рассмотренные выше органы чувств влияют на клев рыб. Поэтому часто можно встретить советы о том, что не следует перебарщивать с ароматизаторами. У человека и у рыб есть определенный порог чувствительности по запахам. При его переходе уже не ощущается ни вкуса, ни запаха. Чтобы понять, как это может быть, достаточно вдохнуть аромат сильного жидкого концентратора с ярко-выраженным запахом. По началу ощущается запах, но потом происходит адаптация. Если вдохнуть жидкость с более сильным запахом, то вы ничего не определите. То же самое происходит и с рыбами. Если забросить прикормку с через чур сильным запахом, то рыба по запаху не отличит ее от грунта на дне.
Зрение у рыб
Нетрудно заметить, что у каждой рыбы имеется пара глаз. Как и у других живых существ, глаза отвечают за зрение. Этот орган по-разному адаптирован к воде у различных видов рыб. Тот же налим практически слепой. Сом также не отличается остротой зрения. А вот та же щука или хариус очень хорошо ориентируются и в цветах, и в оттенках. Для них зрения является одним из основных органов чувств.
Установлено, что рыбы способны хорошо видеть на расстоянии от 5 до 15 метров. Дальше только различаются силуэты. Это связано с особенностями строения хрусталика и сетчатки. Вообще, наблюдается четкая зависимость между величиной глаза и способностью видеть. Чем больше глаз, тем дальше видит их обладатель.
При помощи зрения рыба обнаруживает добычу или хищника. Глаза помогают ей хорошо охотиться, либо вовремя скрываться и не стать легкой добычей. Зрение позволяет определять величину предметов, их форму, а также находить пищу и определять расстояние между объектами.
Что касается способнос
рыб | Определение, виды и факты
Рыба , любой из примерно 34 000 видов позвоночных животных (тип Chordata), обитающих в пресных и соленых водах мира. Живые виды варьируются от примитивных миног и миксин до хрящевых акул, скатов и скатов до многочисленных и разнообразных костистых рыб. Большинство видов рыб хладнокровны; однако один вид, опа ( Lampris guttatus ), является теплокровным.

Тыквенный подсолнух ( Lepomis gibbosus ).
Жак Сикс
Британская викторина
Рыба в море: факт или вымысел?
Может ли рыба испытывать жажду? Акулы используют электричество для охоты? Изучите факты в этой викторине о наших друзьях с плавниками.
Термин рыб применяется к множеству позвоночных нескольких эволюционных линий.Он описывает форму жизни, а не таксономическую группу. Как представители филума Chordata, рыбы имеют общие черты с другими позвоночными. Это жаберные щели в какой-то момент жизненного цикла, хорда или опорный стержень скелета, дорсальный полый нервный шнур и хвост. Живые рыбы представляют собой около пяти классов, которые так же отличаются друг от друга, как и четыре класса знакомых им дышащих воздухом животных: земноводные, рептилии, птицы и млекопитающие. Например, у бесчелюстных рыб (Agnatha) есть жабры в мешочках и отсутствуют пояса конечностей.Сохранившиеся агнатаны - это миноги и микробы. Как следует из названия, скелеты рыб класса Chondrichthyes (из chondr , «хрящ» и ихти, , «рыба») полностью состоят из хряща. У современных рыб этого класса нет плавательного пузыря, а их чешуя и зубы состоят из того же материала плакоидов. Акулы, скаты и скаты - примеры хрящевых рыб. Костистые рыбы - безусловно самый большой класс. Примеры варьируются от крошечного морского конька до голубого марлина весом 450 кг (1000 фунтов), от плоской подошвы и камбалы до квадратной фугу и океанской солнечной рыбы.В отличие от чешуи хрящевых рыб, чешуя костистых рыб, если присутствует, растут на протяжении всей жизни и состоят из тонких перекрывающихся пластинок кости. У костистых рыб также есть жаберная крышка, закрывающая жаберные щели.
Изучение рыб, ихтиология, имеет большое значение. Рыбы представляют интерес для людей по многим причинам, наиболее важными из которых являются их взаимоотношения и зависимость от окружающей среды. Более очевидная причина интереса к рыбам - это их роль как умеренного, но важного компонента мирового продовольственного снабжения.Этот ресурс, который когда-то считался неограниченным, теперь осознается как конечный и находящийся в тонком балансе с биологическими, химическими и физическими факторами водной среды. Чрезмерный вылов рыбы, загрязнение и изменение окружающей среды - главные враги надлежащего управления рыболовством как в пресных водах, так и в океане. (Подробное обсуждение технологии и экономики рыболовства, см. В коммерческом рыболовстве ). Еще одна практическая причина изучения рыб - их использование для борьбы с болезнями.Как хищники на личинках комаров, они помогают бороться с малярией и другими болезнями, передаваемыми комарами.
Рыбы являются ценными лабораторными животными во многих аспектах медицинских и биологических исследований. Например, готовность многих рыб к акклиматизации в неволе позволила биологам изучать поведение, физиологию и даже экологию в относительно естественных условиях. Рыбы сыграли особенно важную роль в изучении поведения животных, где исследования рыб предоставили широкую основу для понимания более гибкого поведения высших позвоночных.Рыба-зебра используется в качестве модели при изучении экспрессии генов.
Получите эксклюзивный доступ к контенту из нашего первого издания 1768 с вашей подпиской. Подпишитесь сегодняИнтерес к рыбам имеет эстетические и рекреационные причины. Миллионы людей держат живых рыб в домашних аквариумах для простого удовольствия наблюдать за красотой и поведением животных, которые в противном случае были бы им незнакомы. Аквариумные рыбки представляют собой личную задачу для многих аквариумистов, позволяя им проверить свою способность удерживать небольшой участок естественной среды в своих домах.Спортивная рыбалка - это еще один способ насладиться природой, которым каждый год балуются миллионы людей. Интерес к аквариумным рыбкам и спортивной рыбалке поддерживает многомиллионные отрасли промышленности по всему миру.
Общие характеристики
Структурное разнообразие
Рыбы существуют уже более 450 миллионов лет, за это время они неоднократно эволюционировали, чтобы вписаться почти во все мыслимые типы водной среды обитания. В некотором смысле наземные позвоночные - это просто сильно модифицированные рыбы: когда рыбы колонизировали среду обитания на суше, они стали четвероногими (четвероногими) наземными позвоночными.Популярное представление о рыбе как о скользком, обтекаемом водном животном, обладающем плавниками и дышащим жабрами, применимо ко многим рыбам, но гораздо больше рыб отклоняются от этого представления, чем подчиняются ему. Например, у многих форм тело удлиненное, у других сильно укороченное; тело у одних уплощено (главным образом у донных рыб) и сжато с боков у многих других; плавники могут быть искусно расширены, образуя замысловатые формы, или они могут быть уменьшены или даже потеряны; положение рта, глаз, ноздрей и жабр широко варьируется.Дышащие воздухом появились в нескольких эволюционных линиях.
Многие рыбы имеют загадочную окраску и форму, максимально приближенную к окружающей среде; другие являются одними из самых ярко окрашенных из всех организмов, с широким диапазоном оттенков, часто поразительной интенсивности, на одном человеке. Сияние пигментов может быть усилено структурой поверхности рыбы, так что кажется, что она почти светится. У ряда несвязанных рыб есть настоящие органы, производящие свет. Многие рыбы способны изменять свою окраску - одни для маскировки, другие - для усиления поведенческих сигналов.
Длина взрослой особи варьируется от менее 10 мм (0,4 дюйма) до более 20 метров (60 футов) и вес от примерно 1,5 грамма (менее 0,06 унции) до многих тысяч килограммов. Некоторые живут в неглубоких термальных источниках при температуре немного выше 42 ° C (100 ° F), другие - в холодных арктических морях с температурой на несколько градусов ниже 0 ° C (32 ° F) или в холодных глубоких водах на глубине более 4000 метров (13100 футов) под водой. поверхность океана. Структурная и, особенно, физиологическая адаптация к жизни в таких крайностях относительно плохо изучена и дает любопытным с научной точки зрения большой стимул для изучения.
.| (Органон обонятельный; Нос) Периферический орган обоняния или орган обоняния состоит из двух частей: внешнего носа , носа , который выступает из центра лица; и внутренняя, носовая полость, , которая разделена перегородкой на правую, и левую носовые камеры. | 1 |
| Внешний нос (Nasus Externus; Внешний нос) —Наружный нос имеет пирамидальную форму, и его верхний угол или корень напрямую соединен с лоб; его свободный угол называется вершиной . Его основание перфорировано двумя эллиптическими отверстиями, ноздри , отделены друг от друга переднезадней перегородкой, columna. Края ноздрей снабжены множеством жестких волосков, или вибриссæ, , которые задерживают прохождение посторонних веществ, переносимых потоком воздуха, предназначенного для дыхания. Боковые поверхности носа своим соединением по средней линии образуют dorsum nasi, , направление которого значительно варьируется у разных людей; верхняя часть спины поддерживается носовыми костями и называется мостом . Боковая поверхность внизу заканчивается округлым возвышением, ala nasi. | 2 |
| Структура. - Каркас внешнего носа состоит из костей и хрящей; он покрыт покровом и выстлан слизистой оболочкой. | 3 |
| Костяной каркас занимает верхнюю часть органа; он состоит из носовых костей и лобных отростков верхней челюстиæ. | 4 |
| хрящевой каркас ( cartilagines nasi ) состоит из пяти больших частей, а именно., хрящ перегородки, , , два боковых, и , два больших крыловых хряща, и несколько более мелких частей, , меньшие хрящи крыльев, (рис. 852, 853, 854). Различные хрящи соединены друг с другом и с костями жесткой волокнистой мембраной. | 5 |
| Хрящ перегородки ( cartilago septi nasi ) имеет несколько четырехугольную форму, по краям толще, чем в центре, и завершает разделение между носовыми полостями спереди.Его передний край, сверху наиболее толстый, соединяется с носовыми костями и непрерывен с передними краями боковых хрящей; ниже он соединен фиброзной тканью с медиальными ножками больших хрящей крыла крыльев. Его задний край соединен с перпендикулярной пластинкой решетчатой кости; ее нижний край с сошником и небными отростками верхней челюстиæ. | 6 |
| Он может быть вытянут назад (особенно у детей) в виде узкого отростка, клиновидного отростка , для некоторого расстояния между сошником и перпендикулярной пластинкой решетчатой кости.Перегородочный хрящ не достигает самой нижней части носовой перегородки. Он образован медиальными ножками больших хрящей крыльев крыльев и кожей; он свободно перемещается и поэтому называется подвижной носовой перегородкой . | 7 |
| Боковой хрящ ( cartilago nasi lateralis; верхний боковой хрящ ) расположен ниже нижнего края носовой кости и имеет уплощенную треугольную форму.Его передний край толще заднего и непрерывен сверху с хрящом перегородки, но отделен от него снизу узкой щелью; его верхний край прикрепляется к носовой кости и лобному отростку верхней челюсти; ее нижний край связан фиброзной тканью с большим хрящом крыла крыльев. | 8 |
| Большой хрящ крыловидной мышцы ( cartilago alaris major; нижний боковой хрящ ) представляет собой тонкую гибкую пластину, расположенную непосредственно под предыдущей и согнутую так, чтобы образовывать медиальная и боковая стенки нариса собственной стороны.Часть, которая формирует медиальную стенку ( crus mediale ), слабо связана с соответствующей частью противоположного хряща, причем оба образуют вместе с утолщенным покровом и прилегающей тканью подвижную носовую перегородку . Часть, которая образует боковую стенку ( crus laterale ), изогнута, чтобы соответствовать крылу носа; он овальный и уплощенный, узкий сзади, где он соединен с передним отростком верхней челюсти жесткой фиброзной мембраной, в которой находятся три или четыре небольших хрящевых пластинки, малых хрящей крыловой кости ( cartilagines alares minores; sesamoid хрящи ).Вверху он соединен фиброзной тканью с латеральным хрящом и передней частью хряща перегородки; ниже он не дотягивает до края нариса, а лава завершается жировой и фиброзной тканью, покрытой кожей. Спереди большие хрящи крыльев крыльев разделены выемкой, соответствующей вершине носа. | 9 |
| | |
| РИС. 852– Хрящи носа. Вид сбоку. (Смотрите увеличенное изображение) | |
| | |
| РИС.853– Хрящи носа, вид снизу. (Смотрите увеличенное изображение) | |
| | |
| РИС. 854– Кости и хрящи перегородки носа. Правая сторона. (См. Увеличенное изображение) | |
| мышц , действующих на внешний нос, были описаны в разделе, посвященном миологии. | 10 |
| Покровы тыльной и боковых сторон носа тонкие и слабо связаны с нижележащими частями; но на кончике и на всей поверхности он более толстый и плотно прилегающий и снабжен большим количеством сальных фолликулов, устья которых обычно очень отчетливы. | 11 |
| артерий наружного носа - крыловидная и перегородочная ветви наружной верхней челюсти, которые снабжают кровью и перегородку; дорсальная и боковые стенки снабжены спинной носовой ветвью глазной и подглазничной ветвью внутренней верхней челюсти. вен оканчиваются передней лицевой и офтальмологической венами. | 12 |
| нервов для мускулов носа происходят от лицевых, в то время как кожа получает ответвления от инфратрохлеарной и назоцилиарной ветвей глазной и подглазничной верхнечелюстной. | 13 |
| Носовая полость (Cavum Nasi; Носовая ямка) - Носовые камеры расположены по обе стороны от срединной плоскости. Они открываются спереди через ноздри и сообщаются сзади через хоану с носовой частью глотки. ноздри представляют собой несколько грушевидных отверстий, каждое размером около 2,5 см. переднезадний и 1,25 см. поперечно в самой широкой части. Хоан - это два овальных отверстия размером 2 штуки каждое.5 см. по вертикали и 1,25 см. в поперечном направлении в хорошо развитом черепе взрослого человека. | 14 |
| Описание костных границ носовых полостей см. На страницах 194 и 195. | 15 |
| Внутри отверстия ноздри имеется небольшое расширение, преддверие , ограничен латерально крыловидной и латеральной ножкой большого хряща крыла, а медиально - медиальной ножкой того же хряща.Он покрыт кожей, содержащей волосы и сальные железы, и простирается в виде небольшого углубления к вершине носа. Каждая носовая полость, выше и позади преддверия, разделена на две части: обонятельная область , , состоящая из верхней носовой раковины и противоположной части перегородки, и дыхательная область , , которая включает остальную часть полости. . | 16 |
| | |
| РИС. 855– Боковая стенка полости носа.(См. Увеличенное изображение) | |
| Боковая стенка (рис. 855, 856). —На боковой стенке находятся верхняя, средняя, и нижняя носовые раковины, и ниже, а сбоку от каждой раковины находится соответствующий носовой проход или проход. Над верхней раковиной находится узкое углубление, клиновидно-этмоидальное углубление, , в которое открывается клиновидная пазуха. Верхний проход - короткий косой проход, проходящий примерно на полпути вдоль верхней границы средней раковины; задние решетчатые клетки открываются в переднюю часть этого прохода. средний проход находится ниже и латеральнее средней раковины и продолжается кпереди в неглубокое углубление, расположенное над вестибюлем и названное атриумом среднего прохода. При поднятии или удалении средней раковины боковая стенка этого прохода полностью раскрывается. На нем находится округлая возвышенность, bulla ethmoidalis, и ниже, а перед ней - изогнутая расселина hiatus semilunaris. | 17 |
| bulla ethmoidalis вызывается выпуклостью средних решетчатых ячеек, которые открываются на ней или непосредственно над ней, а размер буллы зависит от размера содержащихся в ней клеток. | 18 |
| | |
| РИС. 856– Боковая стенка носовой полости; три носовые раковины удалены. (См. Увеличенное изображение) | |
| Полулунный перерыв ограничен снизу острым вогнутым краем крючковидного отростка решетчатой кости и ведет в изогнутый канал, воронку , ограниченную сверху the bulla ethmoidalis и ниже боковой поверхностью крючковидного отростка решетчатой кости.Передние решетчатые клетки открываются в переднюю часть воронки, а это немногим более чем на 50%. субъектов непосредственно переходит в лобно-носовой проток или проход, ведущий из лобной воздушной пазухи; но когда передний конец крючковатого отростка сливается с передней частью буллы, эта непрерывность прерывается, и лобно-носовой проток затем открывается непосредственно в передний конец среднего прохода. | 19 |
| Ниже буллы этмоидальной, частично скрытой нижним концом крючковатого отростка, находится ostium maxillare, или отверстие от верхнечелюстной пазухи; на фронтальном срезе это отверстие располагается у свода пазухи.Дополнительное отверстие пазухи часто находится ниже заднего конца средней носовой раковины. нижний проход находится ниже и латеральнее нижней носовой раковины; носослезный проток открывается в этот проход под прикрытием передней части нижней раковины. | 20 |
| Медиальная стенка (рис.854). - Медиальная стенка или перегородка часто более или менее отклоняется от срединной плоскости, таким образом уменьшая размер одной полости носа и увеличивая размер другой; также иногда присутствуют гребни или шпоры кости, врастающие в ту или иную полость из перегородки.Сразу над резцовым каналом у нижнего края хряща перегородки видно углубление, носо-небное углубление , . В перегородке рядом с этим углублением можно различить крохотное отверстие; он ведет назад в слепой карман, рудиментарный вомероназальный орган Якобсона, , который поддерживается полосой хряща, вомероназальный хрящ. Этот орган хорошо развит у многих низших животных, где он, по-видимому, играет роль в обонянии, поскольку он снабжен ветвями обонятельного нерва и покрыт эпителием, аналогичным эпителию обонятельной области носа. . | 21 |
| Крыша носовой полости узкая из стороны в сторону, за исключением ее задней части, и может быть разделена сзади на клиновидную, решетчатую и лобно-носовую части после кости, которые его образуют. | 22 |
| Пол вогнут из стороны в сторону и почти горизонтален переднезадний; его передние три четверти образованы небным отростком верхней челюсти, а задняя четверть - горизонтальным отростком небной кости.В его переднемедиальной части, непосредственно над резцовым отверстием, иногда можно увидеть небольшое углубление, носо-небное углубление , ; он направлен вниз и вперед и занимает положение канала, который соединял носовую полость с ротовой полостью на ранних этапах жизни плода. | 23 |
| Слизистая оболочка ( mucosa nasi ). —Слизистая оболочка носа выстилает носовые полости и плотно прилегает к надкостнице или надхрящнице.Он непрерывен с кожей через ноздри и со слизистой оболочкой носовой части глотки через хоану. Из носовой полости можно проследить ее непрерывность с конъюнктивой через носослезные и слезные протоки; а также с лобными, решетчатыми, клиновидными и верхнечелюстными пазухами через несколько отверстий в проходах. Слизистая оболочка является самой толстой и наиболее сосудистой над носовой раковиной. Он также толстый над перегородкой; но он очень тонкий в проходах на дне носовых полостей и в различных носовых пазухах. | 24 |
| Из-за толщины большей части этой перепонки носовые полости намного уже, а средняя и нижняя носовые раковины кажутся крупнее и заметнее, чем в скелете; также значительно сужены различные отверстия, сообщающиеся с проходными отверстиями. | 25 |
| Строение слизистой оболочки (рис.857). - Эпителий, покрывающий слизистую оболочку, различается по своему характеру в зависимости от функций той части носа, в которой он находится.В области дыхания оно столбчатое и реснитчатое. Среди столбчатых клеток вкраплены бокаловидные или муциновые клетки, а между их основаниями находятся более мелкие пирамидные клетки. Под эпителием и его базальной мембраной находится фиброзный слой, инфильтрированный лимфатическими тельцами, так что во многих частях образуется диффузная лимфоидная ткань, а под ней почти непрерывный слой малых и более крупных желез, некоторых слизистых и некоторых серозных, протоков которые открываются на поверхности. В обонятельной области слизистая оболочка желтоватого цвета, а эпителиальные клетки столбчатые и не реснитчатые; они бывают двух видов: опорные клетки и обонятельные клетки.Опорные клетки содержат овальные ядра, которые расположены в более глубоких частях клеток и составляют зону овальных ядер; поверхностная часть каждой клетки столбчатая и содержит гранулы желтого пигмента, в то время как ее глубокая часть представляет собой тонкий процесс, который разветвляется и взаимодействует с аналогичными процессами соседних клеток, образуя сеть в слизистой оболочке. Между глубокими отростками опорных клеток находится ряд биполярных нервных клеток, обонятельных клеток, по каждая, состоящих из небольшого количества гранулярной протоплазмы с большим сферическим ядром и обладающих двумя отростками - поверхностным, который проходит между столбчатые эпителиальные клетки и выступают на поверхности слизистой оболочки в виде тонкого волосоподобного отростка, обонятельных волос; другой, или глубокий, отросток идет внутрь, часто состоит из бусинок и продолжается как аксон волокна обонятельного нерва.Под эпителием и сквозь толщу слизистой оболочки находится слой трубчатых, часто разветвленных желез, желез Боумена, идентичных по структуре серозным железам. Эпителиальные клетки носа, зева и дыхательных путей играют важную роль в поддержании равномерной температуры благодаря влажности, которой они всегда слегка смазывают поверхность. | 26 |
| Сосуды и нервы. - артерий. носовых полостей - передняя и задняя решетчатые ветви глазницы, снабжающие решетчатые клетки, лобные пазухи и крышу носа; клиновидно-небная ветвь $$$, которая снабжает слизистую оболочку, покрывающую раковину, проходы и перегородку, перегородочную ветвь верхней губной кости наружной верхней челюсти; подглазничные и альвеолярные ветви внутренней верхней челюсти, кровоснабжающие выстилочную оболочку гайморовой пазухи; и глоточная ветвь той же артерии, расположенная до клиновидной пазухи.Разветвления этих сосудов образуют плотную плексиформную сеть под и внутри слизистой оболочки. | 27 |
| | |
| РИС. 857– Разрез обонятельной слизистой оболочки. (Cadiat.) (См. Увеличенное изображение) | |
| вены образуют тесное кавернозное сплетение под слизистой оболочкой. Это сплетение особенно хорошо заметно над нижней частью перегородки, а также над средней и нижней раковинами.Некоторые из вен открываются в клиновидно-небную вену; другие присоединяются к передней лицевой вене; некоторые сопровождают решетчатые артерии и заканчиваются офтальмологическими венами; и, наконец, некоторые сообщаются с венами на орбитальной поверхности лобной доли мозга через отверстия в решетчатой пластинке решетчатой кости; когда отверстие слепой кишки открытое, оно передает вену в верхний сагиттальный синус. | 28 |
| Лимфатические сосуды уже были описаны (стр.695). | 29 |
| нервов обычного ощущения - это носоцилиарная ветвь глазного дна, нити от передней альвеолярной ветви верхней челюсти, нерв крыловидного канала, носо-небный, передний небный и носовые ветви клиновидно-небного ганглия. | 30 |
| Назоцилиарная ветвь глазного дна распределяет нити к передней части перегородки и боковой стенке носовой полости.Нити переднего альвеолярного нерва снабжают нижний проход и нижние раковины. Нерв крыловидного канала снабжает верхнюю и заднюю части перегородки и верхнюю раковину; и верхние носовые ветви от клиновидно-небного ганглия имеют аналогичное распределение. Носо-небный нерв снабжает середину перегородки. Передний небный нерв снабжает нижние носовые ветви средней и нижней раковинами. | 31 |
| Обонятельный нерв , особый нерв обоняния , распространяется на обонятельную область.Его волокна возникают из биполярных обонятельных клеток и лишены мозговых оболочек. Они объединяются в пучки, которые образуют сплетение под слизистой оболочкой, а затем поднимаются вверх по бороздкам или каналам в решетчатой кости; они проходят в череп через отверстия в решетчатой пластинке решетчатой кости и проникают в нижнюю поверхность обонятельной луковицы, где они разветвляются и образуют синапсы с дендритами митральных клеток (рис. 772). | 32 |
| | |
| РИС.858– Нервы перегородки носа. Правая сторона (см. Увеличенное изображение) | |
| Добавочные пазухи носа (Sinus Paranasales) (рис. 855, 856, 859). | 33 |
| добавочных пазух или воздушных ячеек носа - это лобных, решетчатых, клиновидных, и верхнечелюстных; они различаются по размеру и форме у разных людей и выстланы реснитчатой слизистой оболочкой, непосредственно непрерывной с слизистой оболочкой носа. | 34 |
| Лобные пазухи ( sinus frontales ), расположенные за надбровными дугами, редко бывают симметричными, а перегородка между ними часто отклоняется в ту или иную сторону от средней линии. Их средние размеры следующие: рост 3 см .; ширина 2,5 см .; глубина спереди назад, 2,5 см. Каждый открывается в переднюю часть соответствующего среднего прохода носа через лобно-носовой проток, который пересекает переднюю часть лабиринта решетчатой кости.Отсутствующие при рождении, они обычно довольно хорошо развиты между седьмым и восьмым годами, но достигают своего полного размера только после полового созревания. | 35 |
| | |
| РИС. 859– Венечный разрез носовых полостей. (См. Увеличенное изображение) | |
| Этмоидальные воздушные клетки ( cellulæ ethmoidales ) состоят из многочисленных тонкостенных полостей, расположенных в решетчатом лабиринте и завершенных лобной, верхней, слезной, клиновидной и клиновидной областями. небный.Они лежат между верхними частями носовых полостей и орбитами и отделены от этих полостей тонкой костной пластинойæ. По бокам они разделены на три группы: передних, средних, и задних. Передняя и средняя группы открываются в средний проход носа, первая - через воронку, вторая - на bulla ethmoidalis или выше. Задние клетки открываются в верхний проход под прикрытием верхней носовой раковины; иногда одна или несколько открывается в клиновидную пазуху.Этмоидные клетки начинают развиваться во время жизни плода. | 36 |
| Клиновидных пазух ( sinus sphenoidales ), содержащихся в теле клиновидной кости, различаются по размеру и форме; из-за бокового смещения промежуточной перегородки они редко бывают симметричными. Ниже приведены их средние размеры: высота по вертикали 2,2 см; поперечная ширина 2 см .; переднезадняя глубина 2,2 см. Когда они очень большие, они могут доходить до корней крыловидных отростков или больших крыльев и могут вторгаться в базилярную часть затылочной кости.Каждая пазуха сообщается с клиновидной впадиной через отверстие в верхней части ее передней стенки. При рождении они представлены в виде мелких полостей, но основное их развитие происходит после полового созревания. | 37 |
| | |
| РИС. 860– - Образец от ребенка восьми дней от роду. Путем сагиттального разреза удаляют латеральную часть лобной кости, lamina papyracea решетчатой кости и латеральную часть верхней челюсти - видны sinus maxillaris, cellulæ ethmoidales, передняя и задняя части, infundibulum ethmoidale и примитивный фронтальный синус.(Дэвис.) 141 (Смотрите увеличенное изображение) | |
| | |
| РИС. 861– Образец от ребенка одного года, четырех месяцев и семи дней. Вид сбоку на лобную, решетчатую и верхнечелюстную пазух. (Дэвис.) (См. Увеличенное изображение) | |
| Верхнечелюстная пазуха ( sinus maxillaris; антральный отдел Highmore ), самая большая из придаточных пазух носа, представляет собой пирамидальную полость в теле верхней челюсти.Его основание образовано боковой стенкой носовой полости, а вершина заходит в скуловой отросток. Его крыша или глазничная стенка часто образована инфраорбитальным каналом, а его дно формируется альвеолярным отростком и обычно составляет от 1/2 до 10 мм. ниже уровня пола носа; в пол выступают несколько конических возвышений, соответствующих корням первых и вторых коренных зубов, и в некоторых случаях пол перфорирован одним или несколькими из этих корней. Размер пазухи варьируется в разных черепах и даже на двух сторонах одного черепа.Вместимость взрослого человека варьируется от 9,5 куб. до 20 куб.см, в среднем около 14,75 куб. Следующие измерения относятся к пазухе среднего размера: высота по вертикали напротив первого коренного зуба - 3,75 см; ширина поперечная 2,5 см .; переднезадняя глубина 3 см. В передне-верхней части его основания есть отверстие, через которое он сообщается с нижней частью hiatus semilunaris; второе отверстие часто можно увидеть в перерыве или сразу за ним. Верхнечелюстная пазуха выглядит как неглубокая бороздка на медиальной поверхности кости примерно на четвертом месяце жизни плода, но достигает своего полного размера только после второго прикуса.142 При рождении его размер составляет около 7 мм. в дорсо-вентральном направлении и в двадцать месяцев около 20 мм. 143 | 38 |
| | |
| РИС. 862– Образец ребенка восьми лет, восьми месяцев и одного дня. Вид сбоку на лобную, решетчатую и верхнечелюстную области пазух, боковые части каждой удалены сагиттальными разрезами. Обратите внимание, что фронтальная пазуха образовалась непосредственно от решетчатой воронки. Также обратите внимание на неполные перегородки в верхнечелюстной пазухе.(Дэвис.) (Смотрите увеличенное изображение) | |
обонятельная система | Части, функции и органы
Обонятельная система , телесные структуры, обслуживающие обоняние. Система состоит из носа и носовых полостей, которые в своих верхних частях поддерживают обонятельную слизистую оболочку для восприятия запаха, а в своих нижних частях действуют как дыхательные пути.
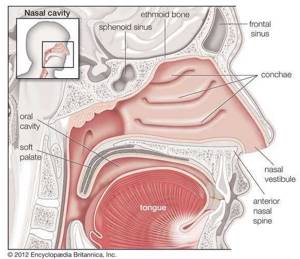 Полость носа человека
Полость носа человека Сагиттальный вид полости носа человека.
Encyclopædia Britannica, Inc.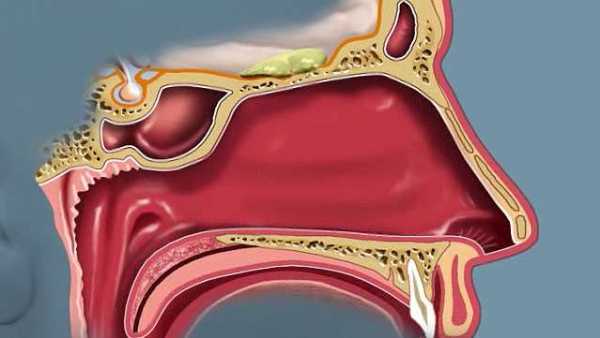 Узнайте, как обонятельная луковица в головном мозге обрабатывает информацию от обонятельных рецепторов носа
Узнайте, как обонятельная луковица в головном мозге обрабатывает информацию от обонятельных рецепторов носа Обонятельная луковица мозга обрабатывает информацию от обонятельных рецепторов носа.
Создано и произведено QA International. © QA International, 2010. Все права защищены. www.qa-international.com Посмотрите все видео по этой статьеКостный каркас носа является частью черепа, но внешний нос поддерживается только костью выше; ниже его форму поддерживают хрящевые пластинки. Расширенная нижняя часть носа, ala, образована только кожей, как снаружи, так и внутри, с фибро-жировой тканью между слоями. Носовые полости разделены перегородкой, на две трети покрытой толстой, сильно сосудистой слизистой оболочкой, состоящей из столбчатого мерцательного эпителия с массами включенных в него ацинозных желез, а в верхней части она покрыта менее сосудистой, но более специализированная обонятельная мембрана.Ближе к передней части нижней части перегородки иногда можно найти небольшое отверстие в короткую глухую трубку, которая идет вверх и назад; это рудиментарный остаток органа Якобсона. Опорный каркас перегородки состоит из решетчатой кости вверху, сошника внизу и перегородочного хряща спереди. Наружная стенка каждой носовой полости разделена на три прохода выступающими носовыми костями. Над верхней носовой костью находится пространство между ней и крышей, известное как recessus sphenoethmoidalis, в задней части которого открывается клиновидный воздушный синус.
 Посмотрите, как исследователи изучают механизм прогнозирующего кодирования, с помощью которого обонятельная система мозга может предугадывать определенные запахи.
Посмотрите, как исследователи изучают механизм прогнозирующего кодирования, с помощью которого обонятельная система мозга может предугадывать определенные запахи. Узнайте, как исследователи исследуют механизм «прогнозирующего кодирования», с помощью которого обонятельная область мозга может предугадывать специфические запахи.
Предоставлено Северо-Западным университетом (издательский партнер Britannica) Просмотреть все видео для этой статьиМежду верхней и средней костями носовых раковин находится верхний проход, который содержит отверстия задних решетчатых воздушных ячеек, а между средней и нижней носовыми раковинами Кости - это средний проход, который является самым большим из трех и содержит округлую возвышенность, bulla ethmoidalis.Сверху и сзади часто есть отверстия для средних решетчатых клеток; внизу и спереди проходит глубокий серповидный желоб, hiatus semilunaris, который сообщается вверху с лобным воздушным синусом, а внизу - с отверстием в антральном отделе Хаймора или антрального отдела верхней челюсти. Нижний проход находится ниже нижней носовой раковины, и, когда он приподнят, видно клапанное отверстие носового протока. Крыша носа узкая, и именно здесь обонятельные нервы проходят через решетчатую пластинку.Пол шире, так что коронковые срезы каждой носовой полости имеют примерно вид прямоугольного треугольника.
Эмбриология
На третьей неделе внутриутробного развития на нижней стороне передней части головы появляются две носовые ямки; они являются первым проявлением истинной обонятельной области носа, и некоторые из их эпителиальных клеток выстилают аксоны, которые ветвятся с дендритами клеток обонятельной доли мозга и таким образом образуют обонятельные нервы.Между обонятельными ямками широкий срединный лобно-носовой отросток растет вниз от области лба, образуя спинку носа и переднюю часть носовой перегородки, а за их пределами латеральные носовые отростки растут вниз и встречаются с верхнечелюстными отростками от первой висцеральной дуги. . Таким образом формируются носовые полости, но они отделены от рта тонкой букконазальной мембраной, которая в конечном итоге прорывается; после этого рот и нос составляют одну полость до формирования неба на третьем месяце.На третьем месяце орган Якобсона можно увидеть как хорошо заметную трубку, выстланную слизистой оболочкой дыхательных путей; Никакого объяснения функции органа Якобсона у людей не известно, и оно, вероятно, полностью атавистично. При рождении носовые полости неглубокие сверху вниз, но быстро углубляются до достижения половой зрелости.
Получите эксклюзивный доступ к контенту из нашего первого издания 1768 с вашей подпиской. Подпишитесь сегодняСравнительная анатомия
В ланцетнике есть ресничная ямка над передним концом центральной нервной системы, которая, вероятно, является рудиментом непарного органа обоняния.У миног ямка сначала вентральная, но позже становится дорсальной и имеет общее отверстие с впячиванием гипофиза. Кроме того, он делится внутри на две боковые половины. У рыб также есть две боковые ямки, ноздри которых выходят иногда, как у акул и скатов, на брюшную поверхность морды, а иногда, как у высших рыб, на спинную поверхность. До этой стадии органы обоняния представляют собой просто ямки, но у иловых рыб от них образуется отверстие в передней части нёба, и поэтому они служат дыхательными путями и органами обоняния.У высших земноводных носовой орган включается в череп, выделяются дыхательная и обонятельная части. В этом классе также обнаруживаются носовые врастания и появляется носослезный проток.
У ящериц обонятельная и дыхательная части очень различны, последняя выстлана только многослойным эпителием, не связанным с обонятельными нервами. Есть одна настоящая носовая кость, растущая из внешней стенки, а рядом с ней находится большая носовая железа. У крокодилов сформировано твердое нёбо, и поэтому между отверстиями наружного и внутреннего ноздрей остается значительное расстояние.У крокодилов также впервые обнаруживаются воздушные пазухи, простирающиеся от обонятельных полостей до костей черепа.
Расположение птиц очень похоже на расположение рептилий; Обонятельная и дыхательная камеры присутствуют, и в последнюю проецируется истинная раковина, хотя есть псевдотурбинальная камера в верхней или обонятельной камере. У млекопитающих обонятельная камера носа развита по-разному: большинство из них макросматичны и имеют большую площадь обонятельной слизистой оболочки; некоторые, такие как тюлени, усатые киты, обезьяны и люди, являются микросматичными, в то время как у зубатых китов обонятельная область практически подавлена у взрослых особей и, как говорят, аносматичны.У макросматических млекопитающих обычно пять костей носовых раковин, хотя у людей их количество меньше. Самый низкий из ряда, челюстно-турбинный, является эквивалентом единственной истинной носовой кости птиц и рептилий и у большинства млекопитающих представляет собой двойной свиток, один лист которого поворачивается вверх, а другой вниз.
Орган Якобсона впервые появляется у земноводных, где он обнаружен как переднезадний желоб в дне носовой полости. У рептилий крыша желоба смыкается с каждой стороны, и формируется трубка, лежащая ниже и внутри носовой полости, открывающаяся кпереди в рот и заканчивающаяся слепой конечностью, кзади от которой проходят ветви обонятельного и тройничного нервов. распределены.У высших рептилий (крокодилы, черепахи и черепахи) и у птиц этот орган подавлен у взрослых. У низших млекопитающих, особенно у однотонных, он очень хорошо развит и заключен в хрящевую оболочку, из которой внутрь ее выступает носовой отросток. У других млекопитающих, за исключением приматов и, возможно, летучих мышей, этот орган весьма обособлен, хотя даже у людей, как было показано, его присутствие может быть продемонстрировано в эмбрионе.
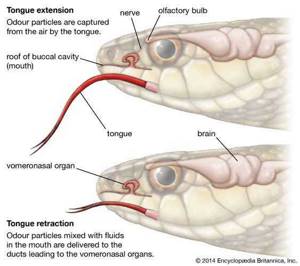 Орган Якобсона; вомероназальный орган
Орган Якобсона; вомероназальный орган Процесс хеморецепции с использованием органа Якобсона, или вомероназального органа.
Encyclopædia Britannica, Inc. .Обзор разработок и приложений
Индексы растительности (VI), полученные на основе данных дистанционного зондирования, представляют собой довольно простые и эффективные алгоритмы для количественной и качественной оценки растительного покрова, силы роста и динамики роста среди других приложений. Эти индексы широко применялись в приложениях RS с использованием различных бортовых и спутниковых платформ, а в последнее время были достигнуты успехи в использовании беспилотных летательных аппаратов (БПЛА). На сегодняшний день не существует единого математического выражения, определяющего все VI, из-за сложности различных комбинаций световых спектров, используемых приборов, платформ и разрешений.Поэтому индивидуальные алгоритмы были разработаны и протестированы в различных приложениях в соответствии с конкретными математическими выражениями, которые объединяют видимое световое излучение, в основном область зеленого спектра, от растительности, и невидимые спектры для получения косвенных количественных оценок поверхности растительности. В реальных приложениях оптимизационные ВП обычно адаптируются к конкретным требованиям приложения вместе с соответствующими инструментами проверки и методологиями на местах. Настоящее исследование знакомит с спектральными характеристиками растительности и обобщает развитие VI, а также преимущества и недостатки различных разработанных индексов.В этом документе рассматривается более 100 ВП, обсуждается их конкретная применимость и репрезентативность в зависимости от интересующей растительности, окружающей среды и точности реализации. Как и ожидалось, исследования и разработки VI, основанных на гиперспектральных платформах и БПЛА, будут иметь широкое применение в различных областях.
1. Введение
Полученная с помощью дистанционного зондирования информация о росте, силе роста и их динамике от наземной растительности может дать чрезвычайно полезную информацию для приложений в мониторинге окружающей среды, сохранении биоразнообразия, сельском хозяйстве, лесном хозяйстве, городской зеленой инфраструктуре и других связанных областях.В частности, эти типы информации, применяемые к сельскому хозяйству, обеспечивают не только объективную основу (в зависимости от разрешения) для макро- и микроменеджмента сельскохозяйственного производства, но также во многих случаях необходимую информацию для оценки урожайности сельскохозяйственных культур [1]. Эти последние приложения были разработаны как хорошо известная категория дисциплин, точное земледелие, начало которой можно было проследить еще три десятилетия назад [1]. Однако применимость дистанционного зондирования и его различных ВП, извлеченных из этих методов, обычно во многом зависит от инструментов и платформ, чтобы определить, какое решение лучше всего подходит для решения конкретной проблемы.
1.1. Соображения относительно платформы дистанционного зондирования
С точки зрения платформ, преимущества спутникового дистанционного зондирования включают высокое пространственное разрешение, которое делает возможным извлечение длинных временных рядов последовательных и сопоставимых данных, что может быть рентабельным [2]. Кроме того, некоторые спутниковые платформы имеют свободный доступ к видимым и многоспектральным данным, например Landsat 7-8. Однако есть две основные проблемы с этими платформами для приложений точного земледелия, которые связаны с разрешением на пиксель (30 м 2 на пиксель для Landsat и 500 м 2 для MODIS) и периодом обращения (16 дней для Landsat и 26 дней для SPOT).Совсем недавно разрешение пикселей было увеличено за счет более новых спутников, таких как WorldView-2 и -3 (DigitalGlobe, Longmont, CO, USA). WorldView-2 был первым коммерческим спутником высокого разрешения, оснащенным восемью спектральными датчиками в видимом и ближнем инфракрасном диапазоне. Наряду с четырьмя типичными мультиспектральными полосами: синим (450–510 нм), зеленым (510–580 нм), красным (630–690 нм) и ближним инфракрасным (NIR) (770–895 нм), каждый датчик имеет узкую фокусировку. в определенном диапазоне электромагнитного спектра, который чувствителен к определенному объекту с земли или свойству атмосферы.Однако изображения с этой платформы могут быть непомерно дорогими для исследований длинных рядов данных.
Вторая проблема спутникового дистанционного зондирования - это время повторного посещения, которое в среднем составляет 16 дней, что затрудняет сельскохозяйственные применения, особенно те, которые связаны с управлением водой и питательными веществами. Более того, пассивные датчики не могут проникать в облака; поэтому для пасмурных дней нет пригодных для использования данных.
Для решения этих двух основных проблем могут использоваться бортовые платформы, а в последнее время и беспилотные летательные аппараты.Первый также может быть непомерно дорогим из-за необходимости в дорогих самолетах и пилотах. Последний стал почти повсеместным за последние пять лет с доступными самолетами и полезными нагрузками камер, начиная от видимого, ближнего и теплового инфракрасного, а также 3D LIDAR, который получил название беспилотной воздушной системы (UAS). Среди платформ БПЛА в основном доступны варианты с фиксированным крылом и мультикоптеры. При использовании этих платформ БПЛА существует компромисс в отношении веса полезной нагрузки и времени полета.В целом, более продолжительное время полета, достигаемое с помощью систем с неподвижным крылом, требует более легкой полезной нагрузки. Например, небольшие камеры видимого диапазона высокой четкости и весом менее 300 граммов в качестве полезной нагрузки БПЛА с неподвижным крылом позволяют ему летать около двух часов, используя доступную в настоящее время батарею [3]. Напротив, мультироторные БПЛА с батарейным питанием и более высокой полезной нагрузкой позволили сократить время полета, которое в настоящее время составляет около 15-25 минут. Используя эти БПЛА, можно достичь более высокого пространственного и временного разрешения данных, что делает возможными прецизионные сельскохозяйственные приложения с субметровым разрешением на пиксель.Это позволяет проводить исследования и практические применения для оценки динамики роста и жизнеспособности растений, определения состояния воды в растениях для составления графиков орошения и моделирования эвапотранспирации, среди прочего [4–9].
1.2. Индексы дистанционного зондирования и растительности
Дистанционное зондирование растительности в основном выполняется путем получения информации об отражении электромагнитных волн от навеса с помощью пассивных датчиков. Хорошо известно, что коэффициент отражения световых спектров от растений изменяется в зависимости от типа растения, содержания воды в тканях и других внутренних факторов [10].Коэффициент отражения от растительности к электромагнитному спектру (спектральная отражательная способность или характеристики излучения растительности) определяется химическими и морфологическими характеристиками поверхности органов или листьев [3]. Основные области применения дистанционного зондирования растительности основаны на следующих спектрах света: (i) ультрафиолетовая область (УФ), которая простирается от 10 до 380 нм; (ii) видимый спектр, состоящий из синей (450–495 нм), зеленой (495–570 нм) и красной (620–750 нм) областей длин волн; 3) ближний и средний инфракрасный диапазон (850–1700 нм) [11, 12].Коэффициент излучения поверхности листьев (эквивалентный поглощающей способности в тепловом диапазоне волн) полностью выросшего зеленого растения без какого-либо биотического или абиотического стресса обычно находится в диапазоне 0,96–0,99, а чаще - между 0,97 и 0,98 [13]. . Напротив, для сухих растений коэффициент излучения обычно имеет больший диапазон от 0,88 до 0,94 [13]. Излучательная способность растений в ближнем и среднем инфракрасном диапазоне широко изучалась в пределах растительного покрова. Индексы, извлеченные из этого диапазона световых спектров, можно отнести к ряду характеристик, помимо количественной оценки роста и жизненной силы растений, связанных, среди прочего, с содержанием воды, пигментов, сахара и углеводов, содержанием белка и ароматических веществ [2, 14].Различные применения зависят от пиков или обертонов отражательной способности для конкретных соединений в видимой и ближней / средней инфракрасной областях светового спектра [14, 15]. Отражательная способность растений в тепловом инфракрасном спектральном диапазоне (8–14 мкм м) подчиняется закону излучения черного тела [16], который позволяет интерпретировать излучение растений как напрямую связанное с температурой растений. Следовательно, индексы, полученные из этого диапазона спектров, могут быть использованы в качестве прокси для оценки динамики устьиц, которая регулирует скорость транспирации растений.Таким образом, более поздние индексы могут использоваться как индикатор состояния воды у растений [17–19] и уровней абиотического / биотического стресса [20, 21].
Последние соображения демонстрируют, что количественная интерпретация данных дистанционного зондирования растительности является сложной задачей. Многие исследования ограничивают эту интерпретацию путем извлечения информации о растительности с использованием отдельных полос светового спектра или группы отдельных полос для анализа данных. Таким образом, исследователи часто объединяют данные из ближнего инфракрасного диапазона (0.7–1,1 м) и красные (0,6–0,7 м) полосы по-разному в зависимости от их конкретных целей [2]. Эти типы комбинаций имеют много недостатков (например, недостаточную чувствительность) за счет использования одной или ограниченной группы полос для обнаружения, например, биомассы растительности. Эти ограничения особенно очевидны при попытке применить эти типы VI на разнородных пологах, таких как садовые плантации деревьев. Смешанное сочетание почв, сорняков, покровных культур в междурядьях и интересующих растений затрудняет распознавание представляющих интерес регионов и извлечение простого VI, особенно когда интересующая растительность имеет разные VI из-за пространственной изменчивости, или VI. соответствующие другой растительности (сорнякам и покровным культурам), которые могут быть похожи на интересующие нас.Последнее усложняет процессы шумоподавления и фильтрации изображений. Для решения этих проблем было разработано несколько методов и алгоритмов анализа изображений, которые будут описаны позже. Несмотря на то, что есть много соображений, описанных ранее, построение простого алгоритма VI во многих случаях могло бы предоставить простые и эффективные инструменты для измерения состояния растительности на поверхности Земли [6].
2. Индексы растительности и процесс проверки
С использованием спектральной аппаратуры высокого разрешения количество полос, получаемых с помощью дистанционного зондирования, увеличивается, а ширина полосы сужается [7].Одним из наиболее часто используемых и реализуемых индексов, рассчитываемых на основе мультиспектральной информации как нормализованное соотношение между красным и ближним инфракрасным диапазонами, является Нормализованный индекс разницы растительности (NDVI) [22]. Непосредственное использование NDVI - это характеристика роста или силы растительного покрова; следовательно, многие исследования сравнивали его с индексом площади листа (LAI) [23], где LAI определяется как площадь односторонних листьев на площадь почвы [24].
Информация о растительности из изображений дистанционного зондирования в основном интерпретируется по отличиям и изменениям зеленых листьев от растений и спектральным характеристикам растительного покрова.Наиболее распространенный процесс проверки - это прямая или косвенная корреляция между полученными ВИ и интересующими характеристиками растительности, измеренными на месте, такими как растительный покров, LAI, биомасса, рост и оценка жизнеспособности. Для оценки ВП используются более устоявшиеся методы с использованием прямых методов и методов с географической привязкой путем мониторинга дозорных растений для сравнения с ВИ, полученными с тех же заводов, для целей калибровки.
Более поздний процесс известен как аллометрические измерения и требует деструктивных методов для сканирования определенной области общего количества листьев на растении или дереве в случае LAI [25].Методы косвенной проверки основаны на проксимальном оборудовании с использованием того же или аналогичного спектрального оборудования для оценки контрольных станций с географической привязкой под тем же углом, что и воздушные платформы. Последний метод полезен для сравнения значений VI, полученных со спутника, которые чувствительны к атмосферным воздействиям и служат средством для получения поправочных коэффициентов. В более поздних косвенных методах, основанных на фотографии покровов, для оценки покрытия растительного покрова, LAI, пористости и индекса комкования использовались автоматизированные методы анализа.
Для этой цели с помощью камер видимого диапазона снимается фотограмметрия крышки, направленная вверх при нулевом зенитном угле, для получения параметров конструкции купола, рассчитанных с использованием алгоритмов компьютерного зрения. Метод автоматического получения и расчета изображений был предложен Fuentes et al. 2008 г. применялся к эвкалиптовым деревьям [26], и он успешно применялся для других культур, таких как виноград, по сравнению с аллометрическими измерениями и для проверки NDVI, рассчитанного на основе спутниковой информации (WorldView-2) [27], яблонь с повышенной точностью с использованием переменной коэффициент ослабления света () [28], и вишневые деревья, улучшающие метод, извлекая нелистовой материал, такой как ветви высоких деревьев [29, 30].В конце 2015 года компьютерное приложение (приложение) для смартфонов и планшетных ПК под названием VitiCanopy было выпущено для бесплатного использования для оценки параметров архитектуры кроны деревьев с использованием автоматизированных алгоритмов фотографирования обложек, которые можно применить к любой другой культуре деревьев, изменив определенное значение [ 31, 32]. Позже были разработаны другие приложения, использующие фотограмметрию RGB для оценки LAI, например PoketLAI [33–35].
2.1. Основные индексы растительности
Иордания [36] предложила в 1969 году один из первых VI под названием Ratio Vegetation Index (RVI), который основан на принципе, согласно которому листья поглощают относительно больше красного, чем инфракрасного света; RVI можно выразить математически следующим образом: где NIR - это коэффициент отражения в ближней инфракрасной области, а - коэффициент отражения в красной полосе.Согласно спектральным характеристикам растительности, кустистые растения имеют низкий коэффициент отражения в красной полосе и показали высокую корреляцию с LAI, сухим веществом биомассы листьев (LDBM) и содержанием хлорофилла в листьях [37]. RVI широко используется для оценки и мониторинга зеленой биомассы, в частности, при высокой плотности растительного покрова, поскольку этот индекс очень чувствителен к растительности и имеет хорошую корреляцию с растительной биомассой. Однако, когда растительный покров разрежен (менее 50%), RVI чувствительны к атмосферным воздействиям, и их представление о биомассе является слабым.
Индекс разницы в растительности (DVI) был предложен позже [38] и может быть выражен как DVI очень чувствителен к изменениям почвенного фона; его можно применять для мониторинга экологической среды растительности. Таким образом, DVI также называют индексом экологической растительности (EVI).
Индекс перпендикулярной растительности (PVI) [38] представляет собой моделирование индекса зеленой растительности (GVI) в данных NIR 2D. В системе координат спектральный отклик от почвы представлен в виде косой черты (линия осветления почвы).Последний эффект можно объяснить тем, что почва дает высокий спектральный отклик в ближнем инфракрасном диапазоне и диапазонах. Расстояние между точкой отражательной способности (, NIR) и линией почвы определено как перпендикуляр VI, который можно выразить следующим образом: где - коэффициент отражения почвы; - коэффициент отражения растительности; PVI характеризует биомассу растительности на почвенном фоне; чем больше расстояние, тем больше биомасса.
PVI можно также количественно выразить как где и - значения яркости отраженного излучения от ближнего ИК-диапазона и, соответственно; - точка пересечения базовой линии почвы и вертикальной оси отражательной способности в ближнем ИК-диапазоне; и θ - угол между горизонтальной осью отражательной способности и базовой линией почвы.Таким образом PVI эффективно отфильтровывает влияние почвенного фона; PVI также имеет меньшую чувствительность к атмосферным воздействиям и в основном используется для инверсии параметров поверхностной растительности (урожайность травы, содержание хлорофилла), расчета LAI, а также идентификации и классификации растительности [39, 40]. Однако PVI чувствителен к яркости и отражательной способности почвы, особенно в случае слабого растительного покрова, и требует корректировки с учетом этого эффекта [41].
Как упоминалось ранее, Нормализованный разностный индекс растительности (NDVI) является наиболее широко используемым в качестве VI; это было предложено Роуз-младшим.и другие. [42], который может быть выражен как: Поскольку индекс рассчитывается с помощью процедуры нормализации, диапазон значений NDVI находится между 0 и 1, что дает чувствительную реакцию на зеленую растительность даже на территориях с низкой растительностью. Этот индекс часто используется в исследованиях, связанных с региональной и глобальной оценкой растительности, и было показано, что он связан не только со структурой и LAI, но и с фотосинтезом растительного покрова [43, 44]. Однако NDVI чувствителен к влиянию яркости почвы, цвета почвы, атмосферы, облаков и теней облаков, а также тени листвы и требует калибровки дистанционного зондирования.
2.2. Индексы растительности с учетом атмосферных воздействий
Учитывая ограничения NDVI при атмосферных воздействиях, Кауфман и Танре [40] предложили Индекс устойчивости к атмосферным воздействиям (ARVI). Этот индекс основан на знании того, что атмосфера оказывает значительное влияние по сравнению с NIR. Таким образом, Кауфман и Танре изменили значение излучения на разницу между синим () и. Следовательно, ОРВИ может эффективно снизить зависимость этого ИВ от атмосферных воздействий, что может быть выражено следующим образом: где - разница между и, - это отражательная способность, связанная с молекулярным рассеянием и газовым поглощением для поправок на озон, и представляет параметры кондиционирования воздуха.
ОРВИ обычно используется для устранения воздействия атмосферных аэрозолей. Эффекты аэрозолей и озона в атмосфере все еще необходимо устранить с помощью модели атмосферного переноса 5S [45]. Однако для реализации модели атмосферного пропускания 5S необходимо учитывать фактические атмосферные параметры, которые трудно получить. Если индекс ARVI не рассчитывается с использованием модели 5S, не ожидается, что этот индекс будет работать намного лучше, чем NDVI, с учетом атмосферных эффектов или крупных пылевых частиц в атмосфере.Таким образом, Zhang et al. [46] предложили новый индекс устойчивости растительности к атмосферным воздействиям, а именно IAVI, который может устранить атмосферные помехи без использования модели 5S. Где диапазон значений γ составляет от 0,65 до 1,12; Значимое значение ? близко к 1 для ОРВИ. После тестирования ошибка, вызванная влиянием атмосферы в IAVI, составляет от 0,4% до 3,7%, что меньше, чем при использовании NDVI в тех же условиях (14–31%).
2.3. Скорректированный индекс растительности почвы
Отличие растительности от почвенного фона было первоначально предложено Ричардсоном и Вигандом [47] путем анализа линии почвы, которую можно рассматривать как линейную зависимость на 2D-плоскости значений спектрального коэффициента отражения почвы между NIR и.Следовательно, его можно рассматривать как исчерпывающее описание большого количества спектральной информации о почве из ряда сред [48]. Многие ВИ, учитывающие влияние почвенного фона, основаны на этом принципе. В дополнение к PVI ((3) - (4)), Индекс атмосферного сопротивления линии почвы (SLRA) был разработан на основе усовершенствованного принципа линии почвы. Затем SLRA был объединен с преобразованным индексом растительности с поправкой на почву (TSAVI) для разработки индекса растительности с атмосферным сопротивлением типа почвы (TSARVI) [49], который будет обсуждаться позже.
Как было показано ранее, NDVI (5) очень чувствителен к фоновым факторам, таким как яркость и оттенок растительного покрова и яркость фона почвы. Исследования показали, что при увеличении яркости фона NDVI также систематически увеличивается. Учитывая влияние почвенного фона, радиация значительно возрастает, когда растительный покров разрежен; и наоборот, снижается БИК-излучение, чтобы сделать связь между растительностью и почвой более ощутимой. Многие VI были разработаны для адаптации к влиянию почвы.
Поскольку NDVI и PVI имеют некоторые недостатки в описании спектрального поведения растительности и почвенного фона, Хуэте [50] установил индекс растительности с поправкой на почву (SAVI), который можно выразить следующим образом:
Вышеупомянутая модель почвы Система растительности была создана для повышения чувствительности NDVI к почвенному фону, где - индекс кондиционирования почвы, который улучшает чувствительность NDVI к почвенному фону. Диапазон составляет от 0 до 1. На практике значения определяются в соответствии с конкретными условиями окружающей среды.Когда степень покрытия растительностью высока, значение близко к 1, показывая, что почвенный фон не влияет на получение информации о растительности. Такого рода идеальные условия редко встречаются в естественной среде и могут быть применимы только в случае большой плотности полога и покрытия [40]. В большинстве обычных условий окружающей среды значение составляет около 0,5. Когда оно близко к 0, значение SAVI равно NDVI. Однако коэффициент должен меняться обратно пропорционально количеству присутствующей растительности, чтобы получить оптимальную поправку на влияние почвы.Таким образом, модифицированный SAVI (MSAVI) заменяет множитель в уравнении SAVI (8) переменной функцией. Таким образом, MSAVI [51] снижает влияние голой почвы на SAVI, которое можно выразить следующим образом:
SAVI намного менее чувствителен, чем RVI (1), к изменениям фона, вызванным цветом почвы или поверхностной почвой. содержание влаги. Три новые версии SAVI (SAVI2, SAVI3 и SAVI4) были разработаны на основе теоретических соображений о влиянии влажных и сухих почв [41]. SAVI2, SAVI3 и SAVI4 снижают эффект яркости фона почвы, принимая во внимание влияние изменения угла падения солнечного света и изменений физической структуры почвы.
Основываясь на реализации MSAVI, Ричардсон и Виганд (1977) предложили модифицированный вторичный индекс растительности с поправкой на почву (MSAVI2) [47], который можно выразить как MSAVI2, не основанный на принципе линии почвы и имеющий более простой алгоритм. ; он в основном используется при анализе роста растений, исследованиях опустынивания, оценке урожайности пастбищ, оценке LAI, анализе органического вещества почвы, мониторинге засухи и анализе эрозии почвы [39].
Baret et al.изучили чувствительность пяти VI (NDVI, SAVI, преобразованный индекс растительности с поправкой на почву (TSAVI), модифицированный индекс растительности с поправкой на почву (MSAVI) и индекс глобального мониторинга окружающей среды (GEMI)) к почвенному фону. Они смоделировали работу виртуальных приборов для различных текстур почвы, уровней влажности и шероховатости, используя модель рассеяния от произвольно наклоненных листьев (ПАРУС). Они определили оптимальное значение SAIL = 0,16 для уменьшения влияния почвенного фона, а затем предложили Оптимизированный индекс растительности с поправкой на почву (OSAVI) [48], который можно выразить следующим образом: где SAIL равен 0.16 и OSAVI не зависит от линии почвы и может эффективно устранять влияние почвенного фона. Однако применение OSAVI невелико; он в основном используется для расчета надземной биомассы, содержания азота в листьях и содержания хлорофилла, среди прочего [52].
2.4. Индекс трансформации зеленоватой шапки кисточкой (GVI, YVI и SBI)
Каут и Томас изучили спектральную картину процесса роста растительности и назвали ее рисунком «колосовидной шапки», включая отражательную способность фона почвы и линию яркости.Преобразование Tasseled Cap - это преобразование исходных полос изображения в новый набор полос с определенными интерпретациями, которые полезны для картирования растительности. Преобразование Tasseled Cap выполняется путем взятия «линейных комбинаций» исходных полос изображения, что по своей концепции аналогично методу многомерного анализа данных, называемому анализом главных компонентов (PCA) [53].
Кепка с кисточкой может конвертировать данные Landsat MSS, Landsat TM и Landsat 7 ETM. Кроме того, для данных Landsat MSS Tasseled Cap выполняет ортогональное преобразование исходных данных, которое преобразует их в четырехмерное пространство.Это преобразование включает индекс яркости почвы (SBI) (14), степень индекса зеленой растительности (GVI) (12) и степень индекса желтой растительности (YVI) (13). Он также включает индекс несоответствия (NSI) в основном для снижения шума. NSI тесно связан с атмосферными эффектами. Для данных Landsat 5 TM результаты Tasseled Cap состоят из трех факторов: яркости, зеленого цвета и третьего компонента, связанного с почвой. Среди них яркость и зеленый цвет эквивалентны SBI и GVI в MSS Tasseled Cap.Третий компонент связан с характеристиками почвы и влажностью. Для данных Landsat 7 ETM преобразование Tasseled Cap Transformation генерирует шесть полос, а именно яркость, зелень, влажность, четвертый компонент (шум), пятый компонент и шестой компонент. GVI, YVI и SBI игнорируют взаимодействие и эффекты атмосферы, почвы и растительности. SBI an
.







